ABBREVIATIONS
ALT, alanine aminotransferase; AMPK, AMP-activated protein kinase; HFFD, high-fat and fructose diet; AST, aminotransferase; AUC, area under the curve; SC-CO2, supercritical CO2; CRV, carvacrol; OREG, oregano; METF, metformin; HDL, high-density lipoprotein; LDL, low-density lipoprotein; MetS, metabolic syndrome; OGTT, oral glucose tolerance test; SeP, selenoprotein P; T2DM, type 2 diabetes mellitus; TG, triglycerides; TNF-α, tumor necrosis factor.
INTRODUCTION
Metabolic syndrome (MetS) encompasses multiple metabolic disorders including abdominal obesity, insulin resistance, glucose intolerance, inflammation, low high-density lipoprotein (HDL) cholesterol levels, high triglyceride (TG) levels, and hypertension [Gunawan et al., 2021]. It is also associated with chronic conditions, such as type 2 diabetes mellitus (T2DM), dyslipidemia, cardiovascular disease, and non-alcoholic fatty liver disease. Its incidence increases with obesity epidemics, affecting about a quarter of the world population and increasing the risk of chronic diseases and creating a serious burden on the health system [Saklayen, 2018]. Although the etiopathogenesis of MetS is not fully understood due to its heterogeneous nature, its short- and long-term complications are important.
Although lifestyle changes and pharmaceutical interventions are generally preferred in MetS treatment, the long-term side effects and high costs of these methods make traditional herbal approaches a complementary alternative important to investigate. Moreover, pharmaceutical interventions are mono-therapeutic and usually target several health outcomes associated with metabolic dysfunction. In this context, medicinal plants containing synergistic and pharmacodynamic bioactive compounds draw attention as therapeutic or preventive agents to manage metabolic disorders [Rodriguez-Casado, 2016].
Carvacrol, one of the main compounds of oregano essential oil, has the potential to be used as a valuable food additive [Ayres Cacciatore et al., 2020; Elbouny et al., 2025]. It exhibits a wide range of biological activities, including anti-inflammatory, immunomodulatory, antioxidative, antitumor, antibacterial, anti-apoptotic, and neuroprotective ones [Sharifi-Rad et al., 2018]. Current research also suggests that it is a compound with therapeutic potential in MetS management [Khalil et al., 2022; Khazdair et al., 2024]. However, although studies on the effects of carvacrol in isolation are increasing, studies examining the effects of an oregano extract as a whole on MetS are limited, and thus more comprehensive evaluations are needed. In this context, some in vivo studies investigating the effects of Origanum species on MetS-related parameters have revealed that oregano extracts may show ameliorative effects on glucose homeostasis, lipid metabolism, and hepatic functions [Abou-Seif & Hozayen, 2023; Elbouny et al., 2025; Lieshchova & Brygadyrenko, 2022]. For example, in rats with obesity and liver damage induced by high-fat diet, Origanum vulgare L. extract decreased body weight, serum aspartate aminotransferase (AST) and alanine aminotransferase (ALT), total cholesterol, TG and low-density lipoprotein (LDL) cholesterol levels, while increasing HDL cholesterol levels [Lee et al., 2024]. In addition, it contributed to the improvement of steatosis by suppressing the expression of genes involved in hepatic lipogenesis. Similarly, in diabetic and hyperlipidemic rats treated with Origanum majorana L. extract, hyperglycemia, hyperinsulinemia, and hepatic adiposity were reported to be markedly reduced [Pasavei et al., 2020]. Although the effects of Origanum onites L. on MetS have not yet been sufficiently studied, a clinical study in individuals with mild hyperlipidemia reported that regular consumption of the aqueous distillate obtained from this species exerted positive effects on the lipid profile, inflammatory markers, and antioxidant enzyme activities [Özdemir et al., 2008]. These data indicates that O. onites extract, which carries the pharmacological effects of carvacrol, is a promising natural agent for the prevention or management of MetS and scientifically supports its investigation in experimental models.
Most previous studies have utilized oregano extracts obtained through conventional extraction techniques, such as hydrodistillation or solvent extraction [Gheitasi et al., 2021; Pasavei et al., 2020], which may involve high solvent consumption, prolonged extraction times, and potential thermal degradation of bioactive compounds [Bitwell et al., 2023]. Additionally, the incomplete removal of organic solvents (such as methanol or hexane) commonly used in traditional extraction can affect the purity of the extract and raise safety concerns. In recent years, green extraction technologies have attracted increasing attention as safer and more sustainable alternatives. Among these, supercritical fluid extraction (SFE) using carbon dioxide (CO2) has emerged as a particularly advantageous method for natural product extraction. SFE exploits the unique properties of supercritical fluids – high density, high diffusivity, low viscosity, and the absence of surface tension – resulting in efficient mass transfer and selective extraction of bioactive compounds. CO2 is the most widely used supercritical fluid due to its non-toxic, non-flammable, and chemically inert nature, wide availability, and relatively mild critical conditions (critical temperature, Tc=31.1°C; critical pressure, Pc=73.8 bar) [Roko et al., 2024]. The extraction process is conducted in a closed system, isolated from air and light, thereby minimizing degradation of sensitive compounds. In addition, CO2 is generally recognized as safe (GRAS), and supercritical CO2 extraction (SC-CO2) yields solvent-free, highly concentrated extracts suitable for applications relevant to both the food and pharmaceutical industries. Although SC-CO2 is inherently non-polar, its solvating power and selectivity can be modulated through the use of co-solvents, enabling efficient extraction of a broad range of phytochemicals [Gregorius et al., 2019; Uwineza & Waśkiewicz, 2020]. Despite these well-documented advantages, the application of SC-CO2 in studies evaluating oregano extracts within the context of the metabolic syndrome remains limited.
Given the above, the present study aimed to investigate the biochemical, physiopathological, and histopathological effects of administering an oregano plant extract obtained through a green extraction method, along with its active compound – carvacrol, in a rat model of metabolic syndrome. Additionally, its strived to elucidate how oregano and carvacrol influence selenoprotein P (SeP) levels in pancreatic and liver tissues and to identify their role in the pathophysiology of MetS.
MATERIALS AND METHODS
Source of plant material
Oregano (O. onites) plant, which was cultivated at the Kırşehir Ahi Evran University, AHIGETAM (Kırşehir Ahi Evran University Traditional and Complementary Medicine Application and Research Centre) Medicinal and Aromatic Plants Application and Research Area (39.137684 N and 34.155483 W, altitude 982), was harvested in its 3rd year on 20 July 2023. The aired parts of O. onites were dried in the shade at ambient temperature and packed in kraft paper to be prepared for analysis.
Extraction of plant material
The dried oregano was ground, and its 150-g portion was placed in a stainless-steel vessel of a supercritical fluid extraction device (F-500, Superex, Konya, Turkey). The sample was fixed between layers of glass wool at the top and bottom of the vessel. The extraction process was carried out fully automatically under 20 MP pressure, 40°C for 3 h, and the CO2 flow rate kept constant at 5 g/min throughout the time. The extract obtained was stored at −20°C to be used in experimental studies. The total extract amount obtained from a single extraction batch was 6.82 g, corresponding to an extraction yield of 4.55 g/100 g dried oregano (w/w). In total, eight extraction batches were prepared for the in vivo experiments, and the extracts were pooled prior to use.
Analysis of phenolic and volatile organic compound profiles
The phenolic compounds of the oregano extract were analyzed using a reverse phase high performance liquid chromatography (RP-HPLC) method reported by Caponio et al. [1999] with modification. Analysis was performed using a system consisting of a 20ACBM system control unit, an SPD-M20A diode array detector, a CTO-10ASVp column oven, an SIL-20ACHT autosampler, and an LC-20AT pump (Shimadzu, Kyoto, Japan). Data were analyzed using LC Solution software (Shimadzu). Chromatographic separations were achieved using a Zorbax C18 column (250×4.6 mm, 5 μm particle size; Agilent Technologies, Santa Clara, CA, USA), and signals from the eluate were monitored at wavelengths of 280 nm and 320 nm. The column temperature was fixed at 30°C. Solvent A (water containing 3% (v/v) formic acid) and solvent B (methanol) were used as a mobile phase by gradient elution. The following elution program was applied at a flow rate of 0.8 mL/min: 93% A + 7% B for 3 min, 72% A + 28% B in 28 min, 67% A + 33% B in 60 min, 58% A + 42% B in 62 min, 50% A + 50% B in 70 min, 30% A + 70% B in 75 min, and 93% A + 7% B in 90 min. The extract was dissolved in the mobile phase and after passing through a 0.45 μm polytetrafluoroethylene filter, 20 μL of a sample solution were injected into the HPLC system. Phenolic compounds were identified by comparing their retention times with those of authentic reference standards. All analytical standards were purchased from Sigma-Aldrich (St. Louis, MO, USA). Quantification was performed using external standard calibration curves constructed at multiple concentration levels. Results were expressed as mg/kg of dry extract.
Volatile organic compounds of the oregano extract administered to the animals were analyzed using a 7890 A gas chromatograph connected to an MSD 5975 C series mass spectrometer (Agilent Technologies) and resolved on a CP WAX 52 CB capillary column (50 m × 0.32 mm ID, 1.2 μm, Agilent Technologies). Helium was used as carrier gas at a flow rate of 1.2 mL/min. Temperature conditions for gas chromatography (GC) were: 60°C initial temperature, 2 min at 60°C, then increasing to 220°C at 2°C/min, after which the temperature was kept constant for 20 min. A 1-μL aliquot was injected in a split mode (20:1). The mass spectrometry (MS) ion source temperature was set at 230°C, and the MS-quadrupole temperature at 150°C. Mass spectra were recorded in the range of 30–500 atomic mass units (amu) and under electron pulse ionization at an electron energy of 70 eV. The identification of the compounds was made by comparing their mass spectra with the Wiley Registry of mass spectral data, 11th edition (John Wiley & Sons, Inc., Hoboken, NJ, USA). Spectrum integrations were performed using MSDCHEM software (Agilent Technologies). Quantification was performed by peak area normalization, and the results were expressed as relative percentages of the total ion chromatogram (TIC).
Animals and experimental design
Wistar albino rats (≈130 g average body weight) were obtained from the Kırşehir Ahi Evran University Experimental Research Centre in Turkey. Before and during the experiment, all animals were housed in rooms with 12 h light and 12 h dark photoperiod and a constant temperature of 22–24°C. The experimental protocol was approved by the Experimental Animals Ethics Committee of Kırşehir Ahi Evran University (Ethics No: 2023/23/4). Prior to the treatment period, animals were gently handled and accustomed to the gavage procedure through a short acclimatization period to minimize stress.
During the experimental period, animals had access to water and feed ad libitum. To induce MetS, the rats were fed a high-fat, high-fructose diet (HFFD). This type of diet was chosen because MetS induced by high-carbohydrate and high-fat dietary mixtures has been shown to better mimic the disease state in humans compared to other methods of inducing the syndrome [Gunawan et al., 2021]. HFFD was formulated based on previously published high-fat diet models, with modifications [Gunawan et al., 2021; Wang et al., 2015]. The diet composition (g/kg diet) was as follows: casein (177 g), l-cystine/dl-methionine (3 g), corn starch (100 g), fructose (262 g), maltodextrin (30 g), cellulose (50 g), butter as a fat source (310 g), corn oil (10 g), mineral mix (10 g), dicalcium phosphate (13 g), calcium carbonate (5.5 g), potassium citrate (16.5 g), vitamin mix (10 g), choline bitartrate (2 g), and food colorant (1 g).
The animals were divided into five groups. In the control group (n=8), the rats were provided with standard rat chow and tap water throughout the experiment. Rats of the MetS group (n=8) were fed a HFFD, which was provided ad libitum as free feeding every day for 10 weeks. Rats of the MetS+CRV group (n=8) were administered HFFD for 6 weeks and at the beginning of the 7th week, 75 mg of carvacrol per kg of body weight was administered by oral gavage every day for 4 more weeks along with HFFD. The dose of carvacrol was determined based on findings from a previous study [Shoorei et al., 2019]. Pure carvacrol (≥98% purity) was purchased from Sigma-Aldrich (St. Louis, MO, USA). The next MetS+OREG group (n=8), was treated with HFFD for 6 weeks, and at the beginning of the 7th week, an oregano extract was administered by oral gavage every day for 4 more weeks at a dose of 500 mg/kg body weight [Raeeszadeh et al., 2024] along with HFFD. In the MetS+METF group (n=8), the rats were fed HFFD for 6 weeks and at the beginning of the 7th week, metformin (Glukofen, Sandoz, Basel, Switzerland) was administered by oral gavage every day for 4 more weeks at a dose of 300 mg/kg body weight [Ajiboye et al., 2016] along with HFFD.
Morphometric analysis of obesity induction
Body weight gain and Lee index were calculated as indicators of obesity. Body weight was measured weekly using an electronic weighing scale. The length of the rats was measured between the nasal and anal region. The recorded measurements were used to calculate the Lee index according to Equation (1):
Blood pressure measurement
Non-invasive tail-cuff blood pressure measurements were performed in rats using a MAY-NIBP250 system (MAY Biotechnology, Ankara, Turkey). Cuffs were routinely checked for patency before experiments. All measurements were conducted in a designated quiet area (22±2°C), and rats were acclimatized for 1 h prior to recording. Animals were gently guided into restraint tubes to minimize movement, with the occlusion cuff placed at the base of the tail and the sensor cuff positioned adjacent to it. A heating chamber maintained at 32°C was used, and rats were warmed for 5 min before and during recordings. Blood pressure was measured by inflating the occlusion cuff to 250 mmHg and deflating over 15 s, while the sensor cuff detected volume changes in the tail. Rats were habituated for at least 7 consecutive days before baseline measurements. For each animal, a total of five measurements were taken at 1-min intervals. The highest and lowest values were excluded, and the mean of the remaining three measurements was used to obtain systolic pressure (SP), diastolic pressure (DP), heart rate, and mean arterial pressure (AP) data. Mean AP was calculated using Equation (2):
Oral glucose tolerance test
Oral glucose tolerance test (OGTT) was performed in rats fasted for 12 h at the end of the experiment. Blood glucose levels of the rats before glucose loading (minute 0) were measured by an ACCU-Check device (Roche Diagnostics, Mannheim, Germany) from blood taken from tail veins. Then, a 50% (w/v) glucose solution was administered to the rats at a dose of 2 g/kg body weight by oral gavage, and blood glucose levels were measured at 30, 60, 90, and 120 min after glucose loading. The area under the curve (AUC) of the OGTT was calculated.
Tissue collection
At the end of the experiment, the animals were fasted overnight, and then the rats from all groups were decapitated under anesthesia with ketamine 75 mg/kg + xylazine 10 mg/kg intraperitoneally. Blood samples collected after decapitation were placed into gold top tubes without ethylenediaminetetraacetic acid (EDTA), and then centrifuged at 5,700×g for 15 min at 4°C to separate the serum. The liver and pancreas were excised, weighed, and then cut into pieces for storage at −80°C. Liver and pancreas tissue samples were also placed in a 10% formaldehyde solution for histological analysis.
Evaluation of gross changes in the liver and adipose tissue
At the end of the experiment, after decapitation, the organs of the rats were excised, excess blood was removed, and weighed on a precision balance and recorded. Additionally, the accumulation of abdominal adipose tissue, including omental, retroperitoneal, and epididymal fat, was observed in situ, and the weighing results were recorded. Results were expressed as absolute organ weights (g).
Biochemical analysis of rat serum and tissue samples
Rat serum TG, LDL, HDL, ALT, and AST were analyzed by an autoanalyzer (Mindray-BS400, Mindray Bio-Medical Electronics Co., Ltd., Shenzhen, China) using commercial kits Ottobc155, Ottobc145, Ottobc144, Ottobc128, and Ottobc127 (Otto Scientific, Otto Scientific, Miami, FL, USA), respectively. SeP and tumor necrosis factor-α (TNF-α) levels in the liver and pancreas of rats were evaluated using enzyme-linked immunosorbent assay (ELISA) kits (Cat. No: 201-11-0765 and 201-11-1158, respectively; SunredBio, Shanghai, China).
Histopathological analysis
Liver and pancreas tissue samples obtained from all rats were initially fixed in a 10% formaldehyde solution for at least 72 h for light microscopic examination. After fixation, the tissue samples were placed in cassettes and washed under running water for 24 h. To remove water, the tissues were dehydrated through a graded ethanol series (50%, 70%, 80%, 90%, 100%). Subsequently, they were cleared in xylene and then embedded in molten paraffin. Sections of 4–5 μm thickness obtained from the prepared paraffin blocks were stained with hematoxylin-eosin (H&E) (Bio-Optica, Milan, Italy; 05-06004/L Harris’ hematoxylin and 05-10002/L eosin Y 1%). The stained sections of liver were assessed for hydropic degeneration and coagulative necrosis in hepatocytes, dilatation in sinusoids, and hyperemia in blood vessels. The pancreatic islets were additionally evaluated for islet histomorphology, cellular changes, and degenerations. In each liver and pancreas section, 10 different areas were scored for each damage parameter, and the average percentage values within the group were calculated. Histopathological changes were graded for liver as follows: 1 (mild) when observed in less than 25% of the tubular epithelium and islet architecture, 2 (moderate) when observed in 25-50%, 3 (severe) when observed in more than 50–75%, and 4 (very severe) when observed in 75–100% (none = 0, mild = 1, moderate = 2, severe = 3, very severe = 4).
Statistical analysis
Statistical analyses were performed using GraphPad Prism version 9.0 (GraphPad Software, San Diego, CA, USA). Data distribution was assessed for normality prior to statistical analysis. Normally distributed data are presented as the mean and standard deviation (SD), whereas non-normally distributed data are presented as the median. For normally distributed data, differences between groups were evaluated using one-way analysis of variance (one-way ANOVA), followed by Tukey’s post hoc test for multiple comparisons. Non-parametric tests, such as the Kruskal–Wallis and Mann–Whitney U tests, were used when the assumption of normality was not satisfied. Differences were considered statistically significant at p<0.05.
RESULTS AND DISCUSSION
Chemical profile of the Origanum onites L. extract
The phenolic compounds of the O. onites extract obtained with the SC-CO2 method were quantitatively determined by HPLC analysis (Table 1). The phenolic acid detected in the highest content was gallic acid (66.01 mg/kg dry extract). This compound is well known as an antioxidant and can significantly contribute to the antioxidant capacity of the extract. Compounds such as caffeic acid and rosmarinic acid were also found at high contents (25.49 and 24.79 mg/kg, respectively). Their anti-inflammatory and antioxidative effects have been widely reported in the literature [Kassa et al., 2021; Xiang et al., 2022]. Rutin (43.09 mg/kg), luteolin (21.28 mg/kg), and apigenin (26.44 mg/kg) were the major flavonoids. These phenolic compounds increase the cell-protective, vascular health-promoting, and antioxidative potential of the extract [Li et al., 2024]. Compounds, such as quercetin, cinnamic acid, vanillic acid, p-coumaric acid, and hesperidin, which were detected in low contents, may act synergistically and enhance biological activity.
Table 1
Content of phenolic compounds in the Origanum onites L. extract obtained by supercritical CO2 extraction.
The volatile organic compound profile of the oregano extract, revealed 59 compounds (Table S1 in Supplementary Materials). They accounted for 9.74% of TIC peak area. The most dominant compound was carvacrol, which accounted for 64.65% of the total volatile organic compounds. This monoterpenic phenolic compound plays an important role in the therapeutic effects of O. onites, including its anti-inflammatory, antioxidative, and metabolic regulatory activities, which have been associated with improvements in metabolic disturbances, such as dyslipidemia, oxidative stress, and inflammation [Khazdair et al., 2024]. Other important volatile compounds of the extract were linalool (10.22%), thymol (4.12%), β-caryophyllene (1.30%), and β-bisabolene (1.36%), i.e., molecules with known anti-inflammatory, antioxidative, and metabolic regulatory effects [Jugreet & Mahomoodally, 2020; Stojanović et al., 2024].
The chromatographic data obtained reveal that SC-CO2 extraction yielded high contents of phenolic, volatile and semivolatile compounds, and that the extract obtained presents a complex compound profile with potential for therapeutic use.
Weight of body, liver, and intra-abdominal adipose tissue in rats
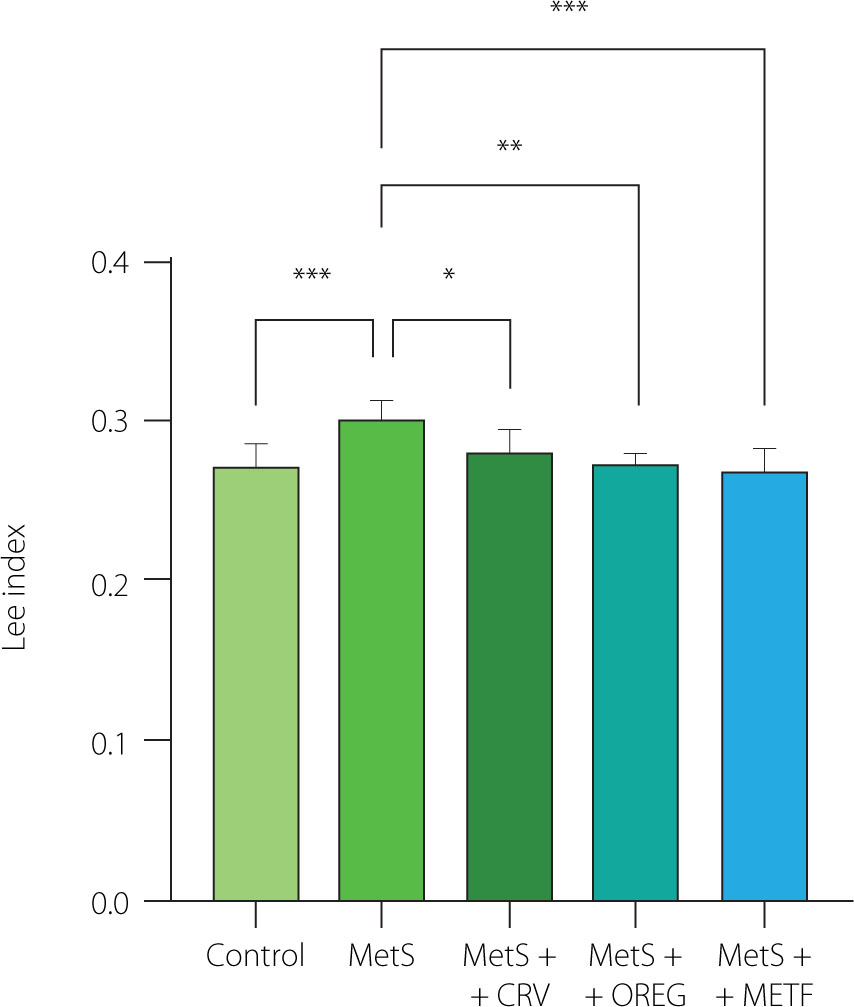
There was no significant difference in the body weight of rats between the groups at the beginning of the experiment (p≥0.05) (Figure 1). At the end of the study (week 10), the body weight of MetS group rats fed the HFFD diet was 375.1 g and was significantly higher (p=0.014) compared to the control group (236.4 g). When the MetS group animals were compared with those of the MetS+METF group, they had a significantly higher body weight (p=0.012). No significant difference was observed in the groups administered the oregano extract and carvacrol compared to the other groups (p≥0.05). The Lee index of the MetS group was significantly higher (p<0.01) compared to the control group (0.30 vs. 0.27) (Figure 2). Moreover, a lower Lee index was computed for the MetS+CRV (0.28), MetS+OREG (0.27), and MetS+METF (0.27) groups compared to the MetS group at p-values of 0.033, 0.020, and <0.01, respectively.
Figure 1
Body weights of rats with induced metabolic syndrome (MetS) and treated with carvacrol (MetS+CRV), oregano extract (MetS+OREG) or metformin (MetS+METF), as well as rats without induced MetS (control) over a 10-week period. Data are represented as mean and standard derivation. Different letters indicate significant differences between the groups in week 10 (p<0.05).
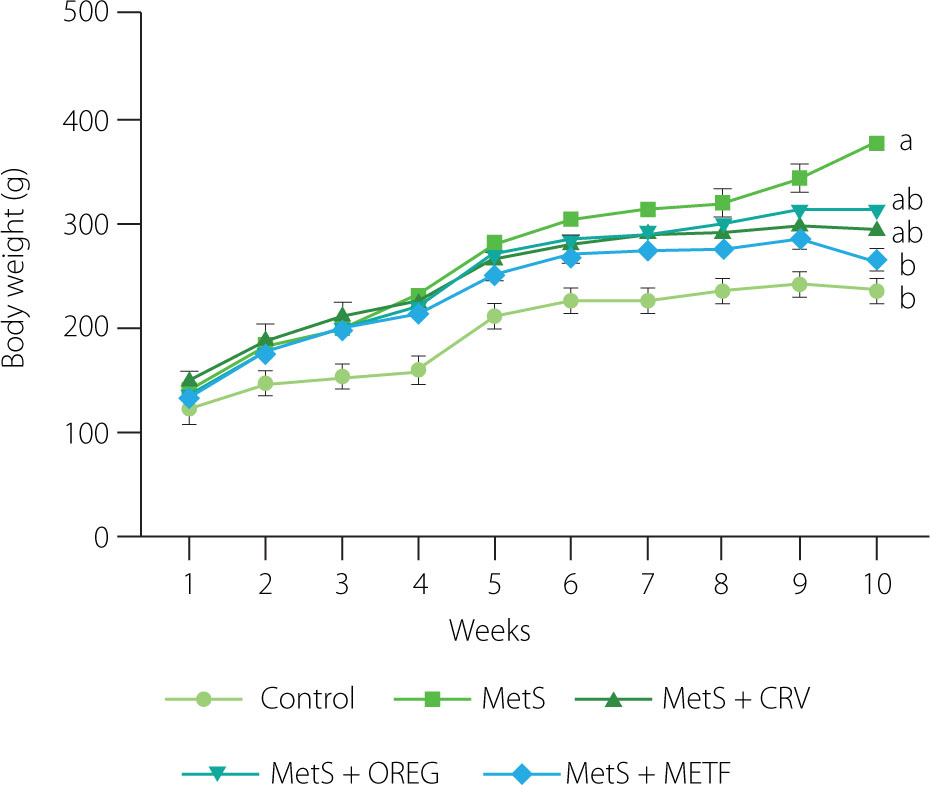
Figure 2
Lee index of rats with induced metabolic syndrome (MetS) and treated with carvacrol (MetS+CRV), oregano extract (MetS+OREG) or metformin (MetS+METF), as well as rats without induced MetS (control) at the end of the 10-week period. Data are represented as mean and standard derivation. Significant differences between groups are indicated with asterisks (*p<0.05, **p<0.01, ***p<0.001).
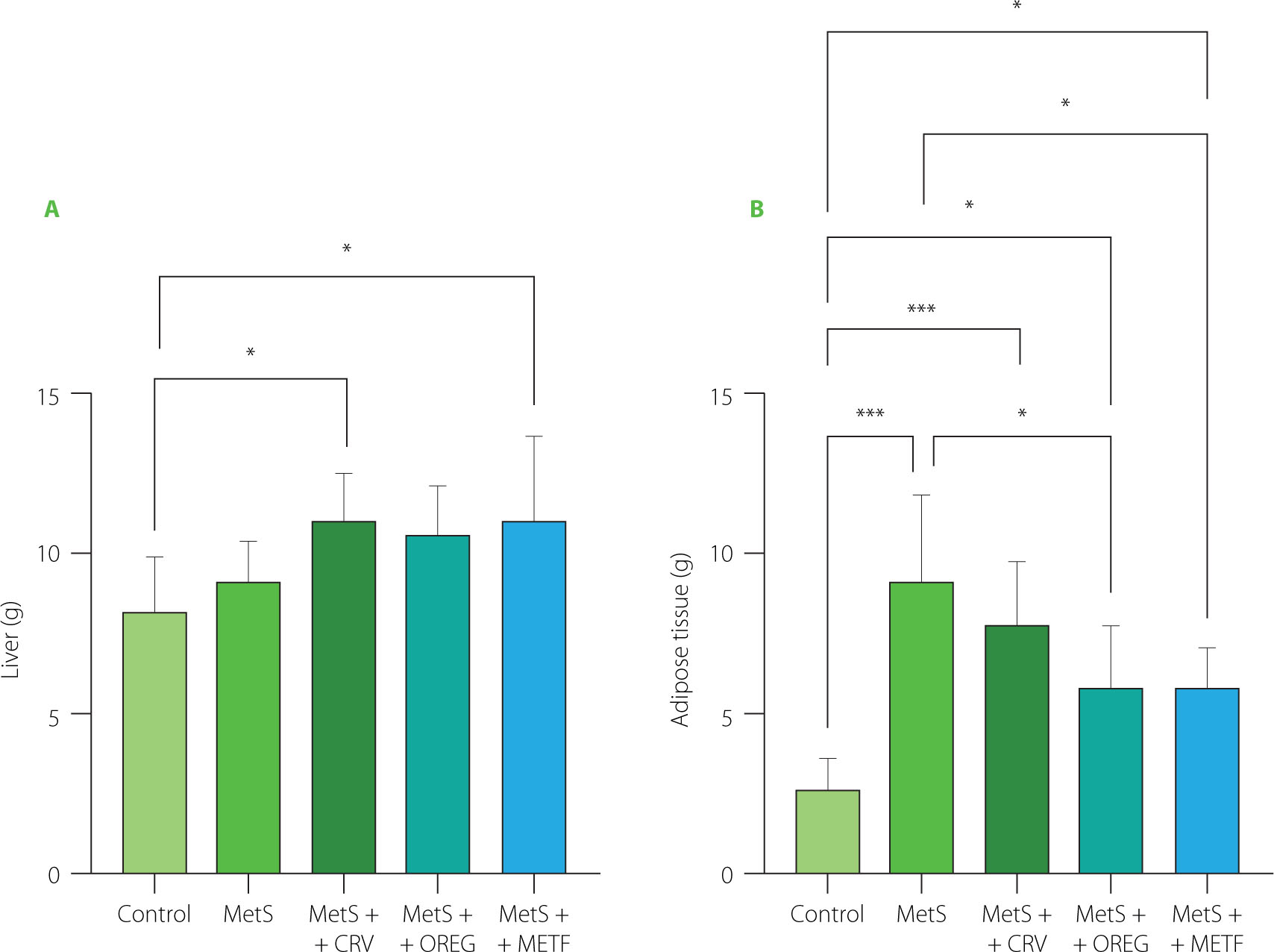
Rat liver and intra-abdominal adipose tissue (including omentum, retroperitoneal, and epididymal adipose tissues) weights are presented in Figure 3. Liver weights were higher in the MetS+CRV group (11.00 g) and the MetS+METF group (11.07 g) compared with the control group (8.17 g) at p=0.045 and p=0.038, respectively. Intra-abdominal adipose tissue analysis also showed that the rats from the MetS group and the MetS+CRV group had significantly higher weights of this tissue when compared to the control animals (9.33 and 7.71 vs. 2.65 g, respectively; p<0.001) Oregano extract treatment significantly lowered the weight of intra-abdominal adipose tissue to 5.86 g compared to the MetS group (p=0.011). Intraabdominal adipose tissue level in the MetS+METF group was also significantly lower (5.86 g) compared to that determined in the MetS group (p=0.022).
Figure 3
Weights of liver (A) and adipose tissue (B) of rats with induced metabolic syndrome (MetS) and treated with carvacrol (MetS+CRV), oregano extract (MetS+OREG) or metformin (MetS+METF), as well as rats without induced MetS (control). Data are represented as mean and standard derivation. Significant differences between groups are indicated with asterisks (*p<0.05, ***p<0.001).
To sum up, the results of this study showed that HFFD administered for 10 weeks promoted obesity parameters that are effective in MetS formation by increasing body weight, liver size, Lee index as well as omental, retroperitoneal, and epididymal fat content. However, oregano extract and carvacrol administration significantly decreased the Lee index of obese rats. Moreover, carvacrol supplementation reduced omental, retroperitoneal, and epididymal fat levels, while oregano extract also led to a significant reduction in these fat depots. Another study demonstrated that oregano seed extract administration reduced weight gain in rats administered a high-fat diet [Lee et al., 2023]. This effect may be due to the presence of phytochemicals that inhibit the pancreatic lipase enzyme by preventing the absorption of dietary fat, thereby increasing fecal excretion of fats and reducing the energy intake of animals. Previous reports have shown that plants rich in phytochemicals, such as saponins, flavonoids, alkaloids and terpenoids, exhibit anti-obesity activity [Rajan et al., 2020]. In the present study, although no significant decrease in body weight was observed among the treatment groups (MetS, MetS+CRV and MetS+OREG), a significant reduction in the Lee index was detected. Since the Lee index is a parameter that takes into account not only body weight but also body length, this finding suggests a favorable change in body composition. The decrease in the Lee index, despite the unchanged body weight, may be attributed to the suppression of visceral adiposity. This suggests the potential of oregano extract and carvacrol to improve body composition without weight loss. Thus, the anti-obesity effects of these compounds may extend beyond weight reduction and involve changes in fat distribution and metabolic profile.
The results indicate that the metabolic syndrome increased liver and intra-abdominal fat accumulation, whereas both oregano extract and metformin treatments mitigated this adverse effect. The observation that oregano extract’s effect was comparable to that of metformin suggests that this herbal treatment may exert potential anti-metabolic or lipid-regulating effects.
Hemodynamic parameters
Analysis of systolic and diastolic blood pressure in rats indicated that values of the MetS group were higher compared to those noted in the control group (Table 2). However, the treatments with oregano extract, carvacrol, and metformin reduced the systolic blood pressure. More precisely, its values were significantly lower in the MetS+CRV, MetS+OREG, and MetS+METF groups when compared to the MetS group. On the contrary, there was no significant effect of oregano extract and carvacrol administrations on diastolic blood pressure in rats. Additionally, heart rate in the rats from the METS group was significantly higher compared to the animals from control group. However, both oregano extract and carvacrol treatments resulted in a reduction of heart rate values.
Table 2
Blood pressure of rats with induced metabolic syndrome (MetS) and treated with carvacrol (MetS+CRV), oregano extract (MetS+OREG) or metformin (MetS+METF), as well as rats without induced MetS (control).
Hypertension is another leading risk factor for morbidity and mortality in MetS. In the case of a high-sugar and high-fat diet, fat cells produce angiotensinogen, which causes an increase in blood pressure [Athyros et al., 2010]. The antihypertensive effects of the oregano extract and carvacrol supplementation observed in this study are in agreement with earlier research findings indicating their potential to decrease blood pressure and support cardiovascular protection by enhancing endothelial function and promoting the synthesis of nitric oxide (NO), a well-known vasodilator [Khazdair et al., 2024; Merimi et al., 2025]. The effects observed in rats with the metabolic syndrome may be partially attributed to free radical scavenging and antihyperglycemic properties of an oregano extract and carvacrol [Cicalău et al., 2021; Radünz et al., 2021].
Blood glucose levels in the oral glucose tolerance test
The OGTT results measured at the end of 10 weeks are shown in Figure 4. At minute 0, the fasting blood glucose level in the rats from the MetS group was significantly higher than that of the control group (p=0.046). Thirty minutes after the administration of a glucose solution at a dose of 2 g/kg body weight, the rats administered the oregano extract (p=0.009), carvacrol (p=0.022), and metformin (p=0.008) showed significantly decreased blood glucose levels compared to the MetS rats. The MetS group had the highest fasting blood glucose value among all groups 120 min after glucose administration. Oregano extract (p=0.002) and carvacrol (p=0.001) supplementation significantly reduced glucose levels at this time point, whereas the reduction observed in the metformin group was not significantly greater (p≥0.05) than that achieved in the MetS+CRV and MetS+OREG groups, despite a lower mean value.
Figure 4
Blood glucose level (A) and area under the curve (AUC) (B) in oral glucose tolerance test of rats with induced metabolic syndrome (MetS) and treated with carvacrol (MetS+CRV), oregano extract (MetS+OREG) or metformin (MetS+METF), as well as rats without induced MetS (control). Data are represented as mean and standar deviation. In graph B, significant differences between groups are indicated with asterisks (**p<0.01, ***p<0.001).
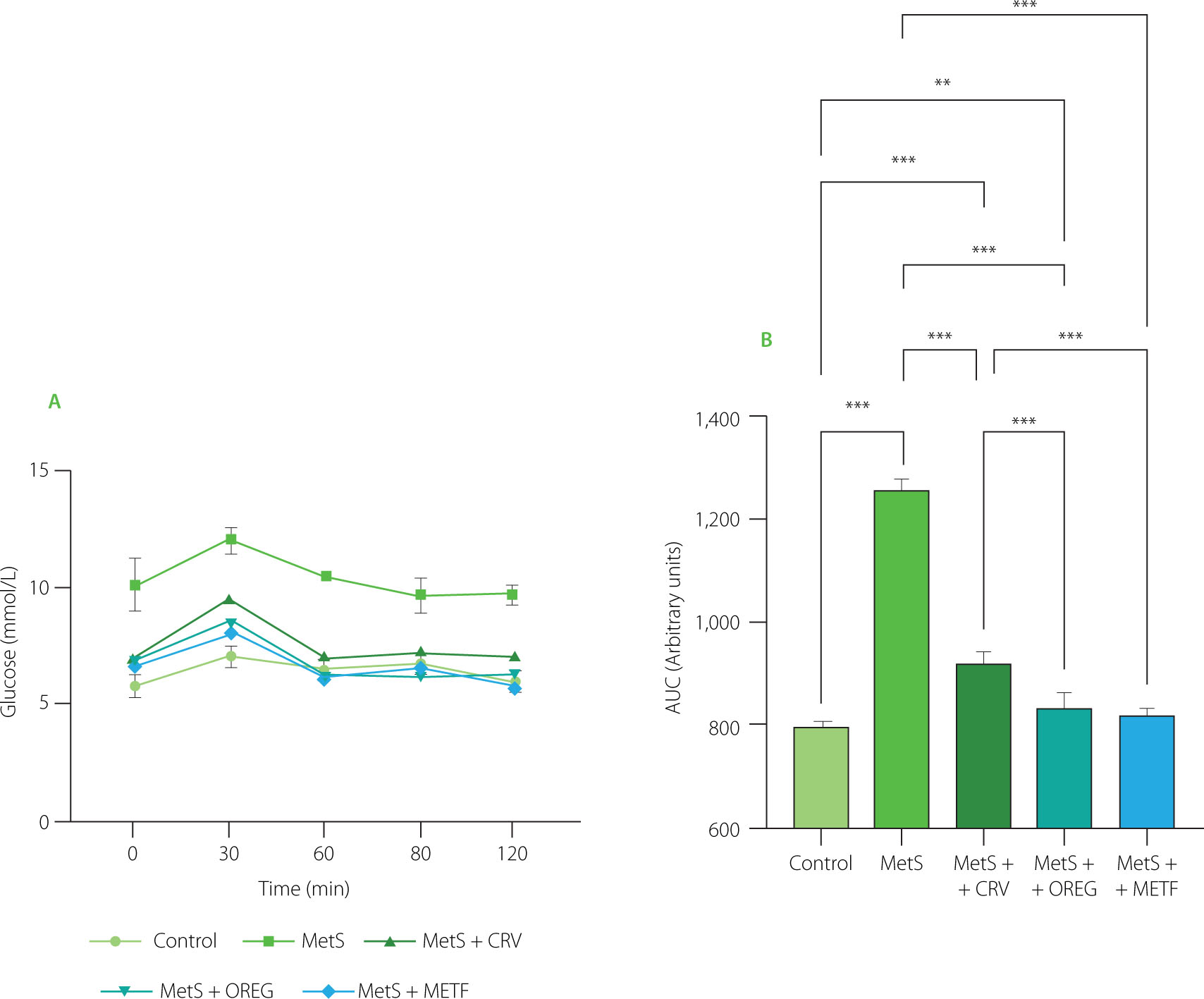
Considering AUC of OGTT, the value for the control group was 789.3, whereas the highest value (1257) was determined for the MetS group (Figure 4). The AUC determined for the combined treatment groups showed lower values, including 919.8 (MetS+CRV), 834.0 (MetS+OREG), and 816.9 (MetS+METF). The results of the Tukey’s multiple comparison test indicated that the AUC of the MetS group was significantly higher than that of the control group (p<0.001). Similarly, AUC determined for MetS+CRV (p<0.001) and MetS+OREG (p=0.003) was significantly higher compared to the control group, whereas the difference between MetS+METF and control was not statistically significant (p=0.097). In turn, the diferences between the MetS group and all combination treatments (MetS+CRV, MetS+OREG, and MetS+METF) were significant at p<0.001. Furthermore, AUC values determined for MetS+OREG and MetS+METF were significantly lower compared to MetS+CRV at p<0.001, while no significant difference was observed between MetS+METF and MetS+OREG (p=0.486).
Another parameter that triggers symptoms in the occurrence of MetS is insulin resistance. Blood glucose levels and AUC values of OGTT performed during the study were significantly increased in the HFFD groups, indicating that feeding HFFD significantly impairs insulin-stimulated glucose uptake in peripheral tissues. This effect may be explained by the fact that a high fat intake reduces the ability of insulin to increase glucose uptake by increasing the release of free fatty acids and fructose that are rapidly metabolized in the liver, increasing TG production [Melo et al., 2021]. Our study results showed that the rats fed a HFFD developed hyperglycemia, which was in agreement with the findings reported by Omidifar et al. [2021]. Furthermore, oregano extract and carvacrol administration decreased the glycemic levels of rats, showing a similar effect as metformin. This healing effect may be due to the presence of phytochemicals that inhibit the digestion of carbohydrates and, therefore, reduce glucose absorption and the postprandial blood sugar level.
Serum biochemical parameters
The findings regarding the effects of carvacrol and oregano extract treatments on serum biochemical parameters are presented in Figure 5. Serum AST levels were significantly (p<0.001) higher in the MetS group, with values of 177.1 U/L compared to 83.79 U/L determined in the control group. ALT values were similar for both groups. Oregano extract and carvacrol treatments reduced AST and ALT values in the metabolic syndrome groups. This was further supported by the significant reductions in AST levels in the MetS+CRV group (122.9 U/L) and MetS+OREG group (107.4 U/L), when compared to the MetS group (p=0.013 and p=0.002, respectively). Similarly, ALT values were significantly lower in the MetS+OREG (49.42 U/L) and MetS+CRV (50.03 U/L) as compared to the MetS group (68.40 U/L) (p=0.033 and p=0.039, respectively). Moreover, in the MetS+METF group, both AST and ALT levels (84.98 and 37.57 U/L, respectively) were significantly higher when compared to the MetS group (p<0.001 and p=0.001, respectively). Similarly, serum TG levels were significantly higher in the MetS group at 173.5 mg/dL compared to the control group at 28.37 mg/dL (p<0.001). At the same time, serum TG levels were significantly reduced in the MetS+CRV group at 30.87 mg/dL, MetS+OREG group at 45.04 mg/dL, and MetS+METF group at 50.24 mg/dL compared to the MetS group (p<0.001). In the evaluation of serum levels of HDL, it was observed that its value was significantly lower in the MetS group when compared with the control group (15.43 vs. 23.70 mg/dL, p=0.004). The values determined in the MetS+CRV (26.32 mg/dL), MetS+OREG (24.88 mg/dL), and MetS+METF (25.60 mg/dL) groups were significantly higher than those assayed in the MetS group (p<0.001, p=0.001, and p<0.001, respectively). Lastly, serum LDL levels were significantly higher in the MetS group (6.24 mg/dL) compared with the control group (3.21 mg/dL) (p=0.008). In the meantime, lower LDL levels were observed in the MetS+CRV (3.69 mg/dL) and MetS+OREG (3.87 mg/dL) groups when compared with MetS group (p<0.05). In this context, the effects of the oregano extract and carvacrol were comparable to those of metformin.
Figure 5
Serum biochemical parameters, including levels of high-density lipoprotein (HDL) cholesterol (A), low-density lipoprotein (LDL) cholesterol (B), triglycerides (TG) (C), aspartate aminotransferase (AST) (D), and alanine aminotransferase (ALT) (E), of rats with induced metabolic syndrome (MetS) and treated with carvacrol (MetS+CRV), oregano extract (MetS+OREG) or metformin (MetS+METF), as well as rats without induced MetS (control). Data are represented as mean and standard deviation. Significant differences between groups are indicated with asterisks (*p<0.05, **p<0.01, ***p<0.001).
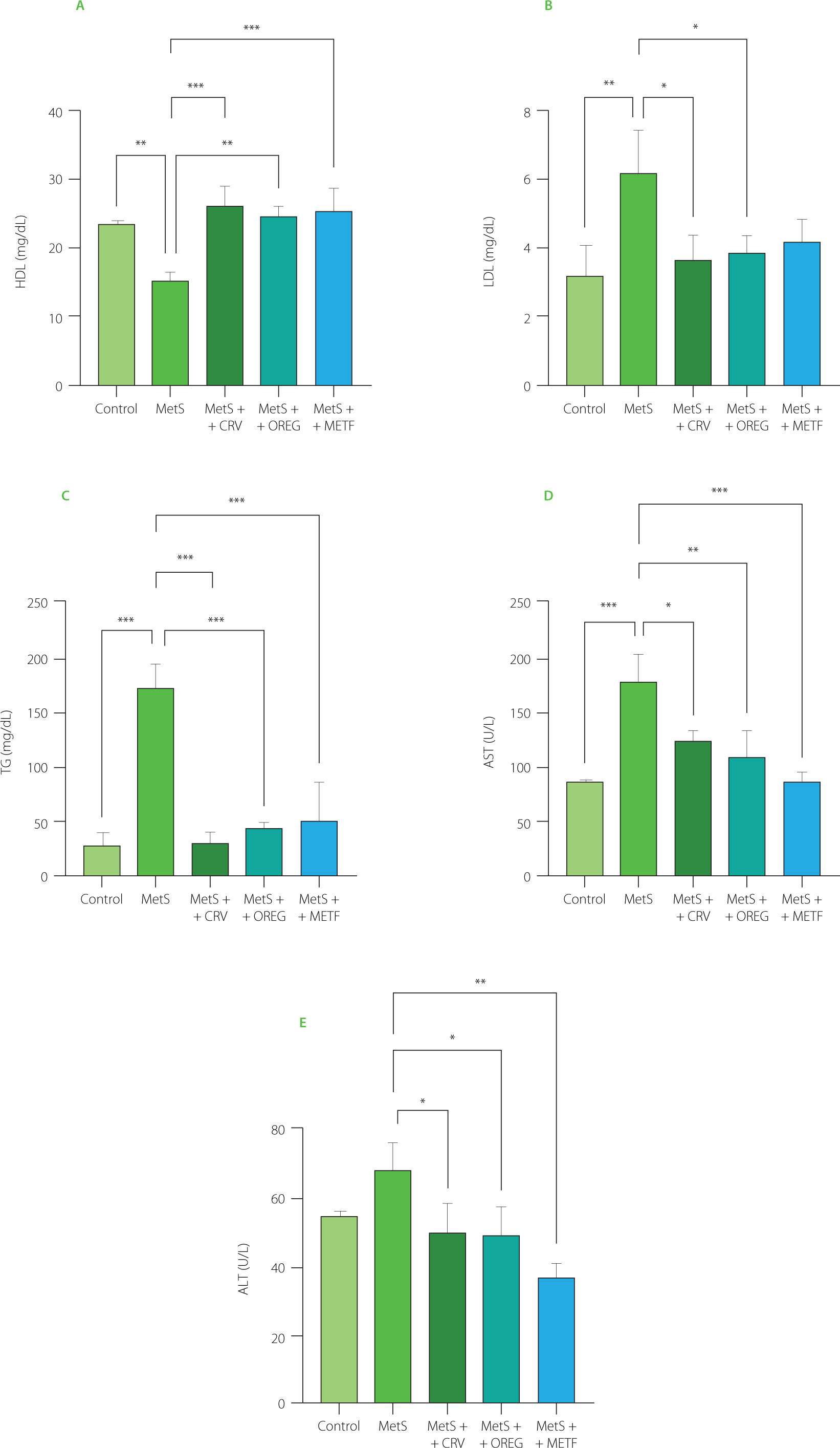
One of the abnormalities that define MetS is dyslipidemia [Gunawan et al., 2021]. In our study, the consumption of the HFFD caused an increase in serum LDL and TG levels, while decreased HDL levels, contributing to a dyslipidemia state. This may be explained by the fact that the high fat content of the diet induces hypercholesterolemia by causing the accumulation of cholesterol plaques in the vascular walls of rats with metabolic disorders [Kenné Toussé et al., 2024]. According to the present study results, oregano extract and carvacrol supplementation decreased TG and LDL levels, while significantly increasing HDL levels. Metformin was less effective than the oregano extract and carvacrol in lowering LDL levels. These effects may be due to the impaired cholesterol synthesis caused by the inhibition of 3-hydroxy-3-methylglutaryl coenzyme A (HMG-CoA) reductase by phytochemicals of the oregano extract. This enzyme is the main regulator of endogenous cholesterol production and its inhibitory effect leads to a decrease in LDL levels [Bampidis et al., 2005]. At the same time, the antioxidative properties of carvacrol prevent free radical damage by reducing lipid peroxidation, which promotes a decrease in TG levels. The increase in HDL level may be attributed to the acceleration of cholesterol transport by increasing apolipoprotein A1 synthesis [Haas et al., 2014]. Thus, oregano and carvacrol positively affect overall cardiovascular health by increasing HDL level while improving both cholesterol and TG levels. Furthermore, the lipid profile results in this study are in agreement with the findings of Özdemir et al. [2008] and Lee et al. [2024], who found that an aqueous distillate of O. onites and an ethyl acetate fraction of O. vulgare seed, respectively, have significant hypocholesterolemic and hypotriglyceridemic effects. Impaired glucose uptake in the liver and muscle tissues leads to insulin resistance and hyperglycemia, triggering hepatic steatosis and inflammatory processes in the liver. This leads to increased lipid peroxidation and oxidative stress in hepatocytes, resulting in elevated ALT and AST levels, which are liver enzymes. Elevated ALT and AST values have been reported in MetS [Nderitu et al., 2023]. This study showed that ALT and AST values were increased in HFFD-fed rats compared to the other treatment groups. Along with the increases in serum ALT and AST, liver histopathological findings from this study showed that HFFD administration caused liver damage. However, in the groups treated with the oregano extract and carvacrol, ALT and AST levels significantly decreased, approximating the control values, and also prevented liver damage. This shows that oregano and carvacrol consumption is safe. These findings are consistent with previous studies reporting hepatoprotective effects of O. vulgare extract, which significantly decreased serum AST and ALT levels in experimental models of liver injury [Habibi et al., 2015].
Liver and pancreas TNF-α levels
TNF-α levels of liver and pancreas tissues are presented in Figure 6. The MetS group had significantly higher hepatic TNF-α levels (119.0 ng/L) when compared to the control group (69.36 ng/L) (p<0.001). In a similar manner, pancreatic levels of TNF-α were significantly higher in the MetS group when compared to the control group; 34.32 vs. 12.29 ng/L, respectively (p<0.001). Carvacrol and oregano extract treatment reduced liver TNF-α level significantly in the MetS+CRV (69.07 ng/L) and MetS+OREG (82.22 ng/L) groups compared to the MetS group (p<0.001). Remarkably, carvacrol drastically decreased pancreatic TNF-α levels, and in the MetS+CRV group, the level reached 20.05 ng/L, which was lower than that in the MetS group (p=0.025). In turn, the oregano extract did not show this kind of activity in terms of pancreatic TNF-α levels. Furthermore, in the MetS+METF group, the liver TNF-α level was significantly lower, measured as 69.25 ng/L, compared to the MetS group. However, the pancreatic level of TNF-α in the MetS+MetF group was significantly raised as compared with the control group, indicating an interaction between metformin and TNF-α modulation in pancreatic tissues.
Figure 6
Tumor necrosis factor-α (TNF-α) levels in the liver (A) and pancreas (B) of rats with induced metabolic syndrome (MetS) and treated with carvacrol (MetS+CRV), oregano extract (MetS+OREG) or metformin (MetS+METF), as well as rats without induced MetS (control). Data expressed as mean and standard deviation. Significant differences between groups are indicated with asterisks (*p<0.05, **p<0.01, ***p<0.001).
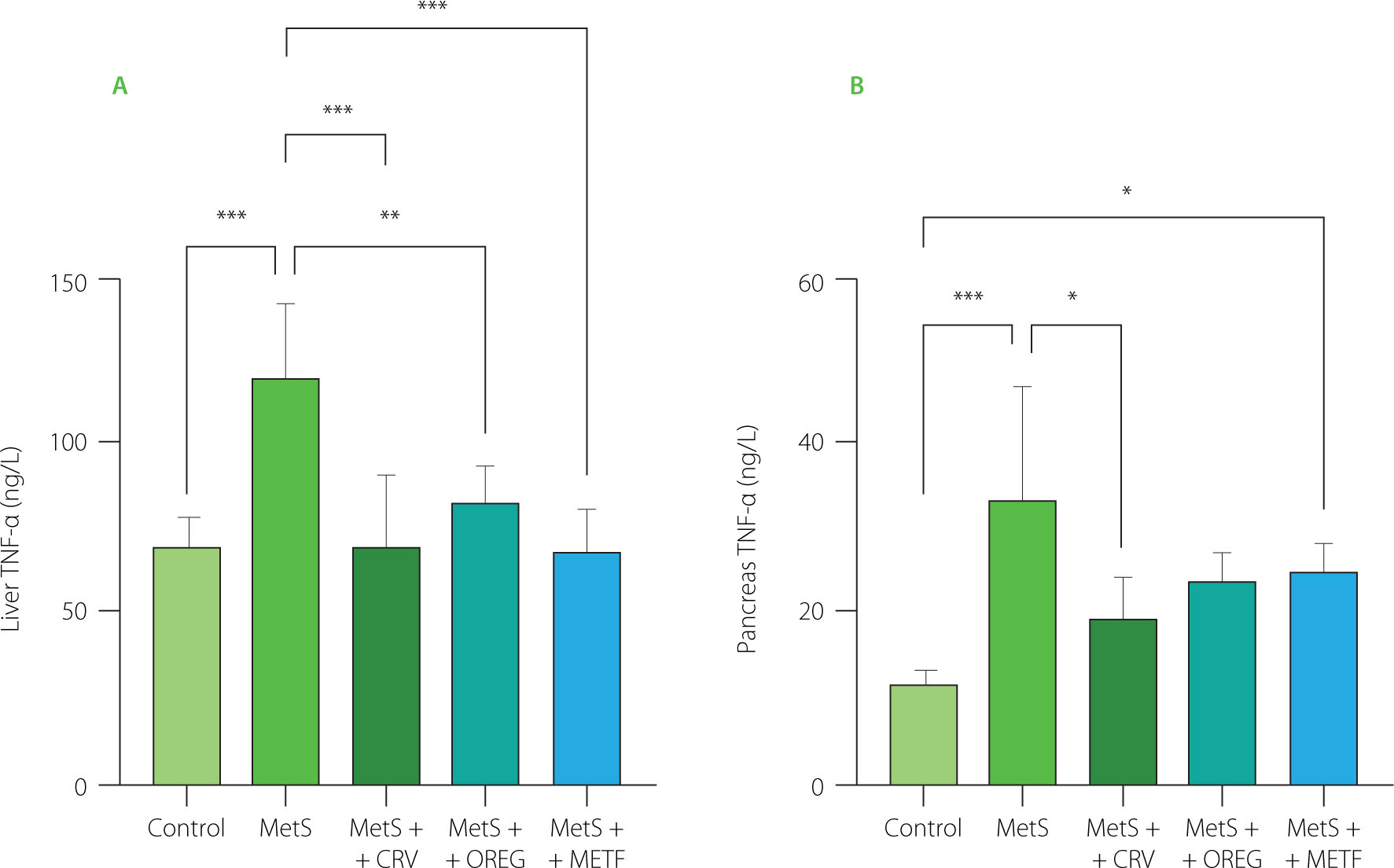
Increased adipose tissue in rats with MetS, correlated with increased macrophages, can trigger the release of various cytokines, such as TNF-α, IL-6 and HMGB1, by switching from the anti-inflammatory ‘M2’ (alternatively activated) to the pro-inflammatory ‘M1’ (classical activation) state [Frisardi et al., 2021]. Overexpression of TNF-α suggests that TNF-α may be a predictor for early diagnosis of MetS [Chen et al., 2021]. In this study, an increase in TNF-α levels was observed in pancreatic and liver tissues of the rats fed a high fat and sugar diet. The oregano extract decreased liver TNF-α level, while carvacrol significantly decreased pancreatic TNF-α level (Figure 6). Phenolic compounds have been reported to exhibit different biological activities, such as antioxidant, anti-inflammatory, antitumoral, and antimicrobial ones [Zhang et al., 2014]. Therefore, the anti-inflammatory effects obtained in this study may be related to the phenolic compounds of the oregano extract. These findings are consistent with the study reported by Sharifi-Rigi et al. [2019] indicating that the O. vulgare extract exerted inhibitory effects on paraquat-induced liver injury due to its antioxidative properties in a rat model of liver injury.
Liver and pancreas selenoprotein P levels
SeP levels of liver and pancreas tissues are presented in Figure 7. The hepatic SeP level in the MetS group was significantly increased to 227.1 ng/mL, compared to the control group showing a value of 144.3 ng/mL (p=0.022). In addition, a significant difference (p=0.016) was observed between the MetS group (227.1 ng/mL) and the MetS+METF group (144.3 ng/mL). In contrast, no significant differences (p≥0.05) were noted in SeP levels among the groups in pancreatic tissue. Most notably, oregano extract and carvacrol had no significant impact on SeP levels in liver and pancreas, indicating that neither of these compounds affects selenium metabolism in MetS.
Figure 7
Selenoprotein P (SeP) levels in the liver (A) and pancreas (B) of rats with induced metabolic syndrome (MetS) and treated with carvacrol (MetS+CRV), oregano extract (MetS+OREG) or metformin (MetS+METF), as well as rats without induced MetS (control). Data expressed as mean and standard deviation. Significant differences between groups are indicated with asterisks (*p<0.05).
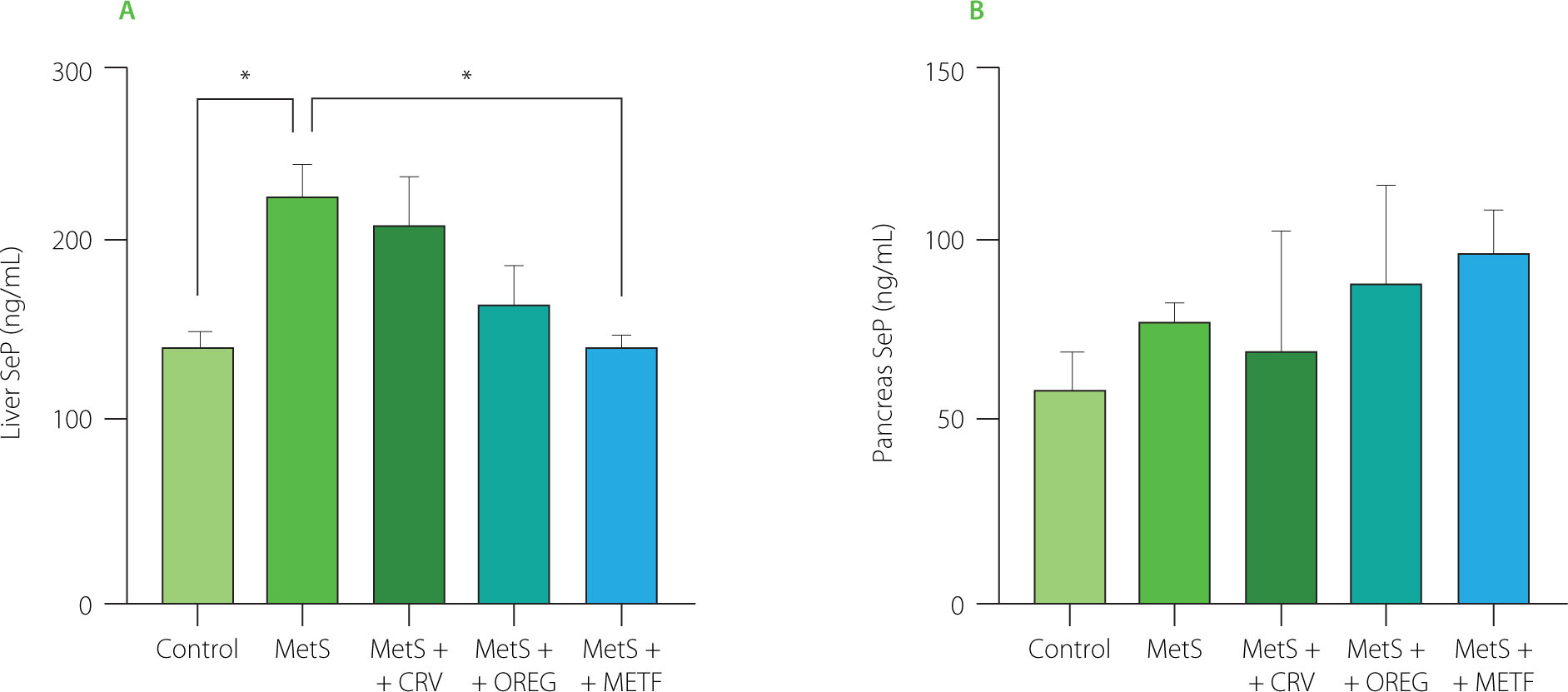
In the present study, SeP tissue levels in liver and pancreas were evaluated to examine the role of SeP in the development of MetS. SeP exhibits antioxidative properties in various tissues as well as acts as an Se transport protein that provides Se to tissues in the body and aids biosynthesis of intracellular selenoproteins [Jensen-Cody & Potthoff, 2021]. No study was found in which SeP levels were analyzed in liver and pancreatic tissues of MetS-induced experimental animals. It has been reported that hepatic expression of SeP and circulating selenium and SeP levels are decreased in rats with acute phase reaction; this impairs hepatic SeP expression during acute phase reaction and thus affects the transport of Se to peripheral tissues [Renko et al., 2009]. In contrast, hepatic SeP levels have been reported to increase with hepatic steatosis and oxidative stress in rats fed a high-fat diet; this change may represent a mechanism against high-fat diet-induced steatosis, oxidative stress, and subsequent inflammation [Murano et al., 2018].
The present study results further showed that there was a significant increase in SeP levels in the liver and no significant increase in pancreatic tissue of the MetS group rats fed a high-fat and high-sugar diet. This finding suggests that SeP may have a regulatory role in pathological processes associated with MetS and may lead to adverse physiopathological processes on liver functions. In our study, oregano extract and carvacrol supplements administered exogenously did not significantly alter SeP levels. However, these supplements have been found to cause a decrease in TG levels. A previous study reported that SeP was positively correlated with plasma TG levels, indicating its association with dyslipidemia [Chen et al., 2017]. This finding suggests that these therapeutic approaches may be effective on lipid profile and inflammation rather than affect SeP metabolism. Moreover, the association of SeP with obesity is another important factor to be considered in the pathogenesis of MetS. In the present study, when body weight changes were analyzed between groups, insulin resistance occurred with increasing body weight in the MetS group and SeP levels increased significantly.
Misu et al. [2010] reported that SeP knockout mice fed a high fat/high sucrose diet were protected against the development of insulin resistance. In addition, SeP decreased AMP-activated protein kinase (AMPK) phosphorylation in the liver, resulting in enhanced fatty acid biosynthesis. This suggests that serum SeP level may exert autocrine effects in the liver. The authors also reported a high glucose-induced up-regulation of SeP in hepatocytes [Misu et al., 2010]. Considering literature data, our study findings support the hypothesis that SeP plays a regulatory role in glucose homeostasis and is also involved in liver physiological processes. Speckmann et al. [2009] reported that treatment with metformin, a drug used in the treatment of diabetes, reduced hepatic SeP mRNA expression and secretion, but the dose of metformin administered was higher than the therapeutic dose in humans (2 mM). According to our study findings, metformin administration decreased the SeP level in the liver of rats with the metabolic syndrome, whereas there was no significant difference in pancreatic tissue. These findings suggest that metformin may exert an effect on liver function through the modulation of SeP levels. However, oregano extract and carvacrol treatments did not produce significant changes in SeP levels, suggesting that their effects may occur through mechanisms other than SeP regulation.
Histopathological assessments
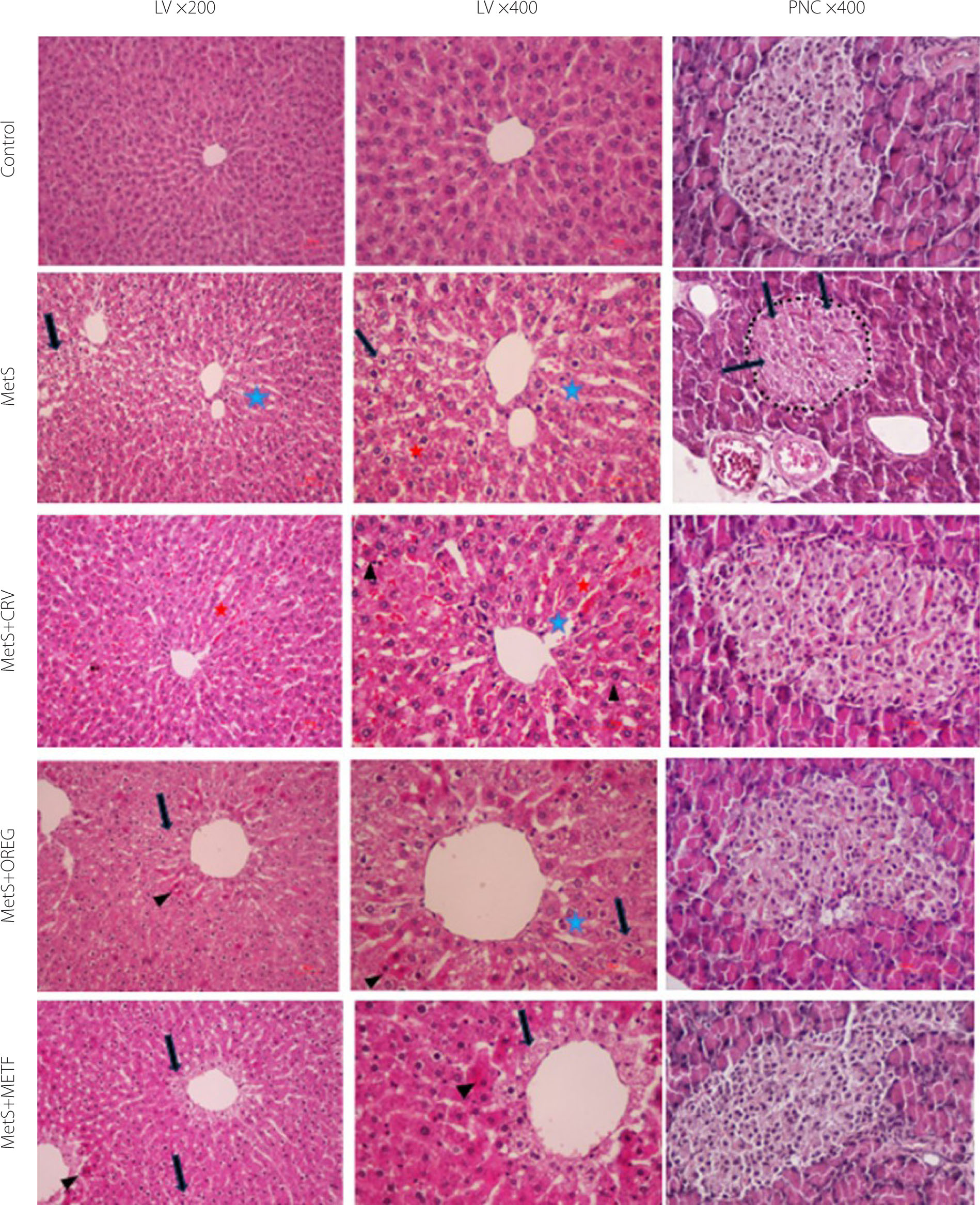
The histopathological images of the liver and pancreas sections of rats from different groups are shown in Figure 8. Furthermore, the results of histopathological scoring of the liver and pancreas are presented in Tables S2 and S3 in the Supplementary Materials, respectively. In the liver sections of the control group, hepatocytes were regularly arranged around the central vein with distinct nuclei, forming orderly cords. The sinusoids between the cell cords exhibited a normal appearance. In the MetS group, where metabolic syndrome was induced by feeding with a specific diet, liver sections showed hydropic degeneration and coagulative necrosis in the parenchyma, particularly in the hepatocytes. There was also irregularity, dilation, and hyperemia in the sinusoids, with disruption of the integrity of the central vein when compared to the control group. In the MetS+CRV group, hepatocytes and cord formations were similar to those of the control group; however, sinusoids were slightly dilated, and there was less coagulative necrosis in the hepatocytes and increased hyperemia. In the MetS+OREG group, liver sections showed minimal hydropic degeneration and coagulative necrosis in hepatocytes in the periacinar region, and less dilation of the sinusoids. Pathological changes were less pronounced in these two groups. In the MetS+METF group, the histomorphology was similar to the control group, although prominent hydropic degeneration was observed in hepatocytes surrounding the central vein in the periacinar region. There was also minimal coagulative necrosis in hepatocytes and reduced sinusoidal dilation. Upon examining the pancreatic sections, the normal islet histomorphology in the control group was disrupted in the MetS groups, showing a scattered and irregular appearance. The cell boundaries became indistinct, and structural abnormalities due to vacuolization, along with hydropic degeneration, were observed. The islets had shrunk, and their boundaries were more blurred. Additionally, a reduction was noted in the number of islet cells. Among the treatment groups, the MetS+METF group showed the most similar appearance to the control group. In the MetS+CRV group, significant improvement was observed compared to the MetS group. The islet contours were distinct and regular, and the cells showed structural and numerical similarities to those in the control group. The MetS+OREG group’s islet cells resembled those in the control group. However, unlike the control group, the number of cells was low, and the islet boundaries were somewhat irregular (Table S2 and S3). These results suggest that the metabolic syndrome induces significant histopathological damage in both hepatic and pancreatic tissues. However, treatments with the oregano extract and metformin alleviate these alterations to varying extents, with metformin showing the strongest protective effect in the pancreas, and oregano extract demonstrating comparable hepatoprotective potential.
Figure 8
Sections of livers (LV) and pancreas (PNC) in different groups of rats. Control group; normal histology in control group. MetS group (LV); coagulation necrosis (arrowheads), severe hydropic degeneration of hepatocytes (black arrow) in the central region, sinusoidal dilatation (blue asterisks) and hyperemia (red asterisks). MetS+CRV group (LV); near-normal histology, comparable to control, but minimal coagulation necrosis, dilatation and hyperemia of the sinusoid. MetS+OREG group (LV); minimal hydropic degeneration and coagulation necrosis of hepatocytes in the periacinar region, mild dilatation of the sinusoids. MetS+METF group (LV); hydropic degeneration and coagulation necrosis in the periacinar region. MetS group (PNC); severe hydropic degeneration of islet cell (black arrow), shrunken and irregular islet borders (a dotted line). MetS+CRV group (PNC); near-normal histology, comparable to control. MetS+OREG group (PNC); near-normal histology, with minimal irregularities in islet borders. MetS+METF group (PNC); near-normal histology.
CONCLUSIONS
This study demonstrates that both SC-CO2–derived O. onites extract and pure carvacrol exert beneficial effects on histological, biochemical, and metabolic alterations in a high-fat, high-fructose diet–induced rat model of MetS. Chemical characterization confirmed that the oregano extract was rich in carvacrol and other bioactive compounds. Treatment with the oregano extract and carvacrol was associated with improvements in lipid parameters, particularly triglyceride levels, and with amelioration of histopathological alterations observed in liver and pancreatic tissues. In addition, both treatments showed anti-inflammatory potential through modulation of TNF-α levels, while hepatic SeP alterations indicated a possible role in metabolic regulation during MetS development.
While the present results support the potential of the O. onites extract and carvacrol as candidates for further investigation in MetS, additional studies focusing on molecular mechanisms, as well as well-designed clinical trials, are required to confirm their translational relevance.