INTRODUCTION
Vegan food alternatives to conventional dairy products present both nutritional advantages and limitations, as reported in multiple studies. Plant-based dairy alternatives (PBDAs) generally contain higher levels of dietary fibre and lower amounts of saturated fat than traditional dairy products, which may contribute to an improved overall diet quality and a reduced saturated fat intake [Marchese et al., 2024; Moshtaghian et al., 2024]. However, most PBDAs tend to provide a lower protein content, with the notable exception of soy-based products, which can match or exceed the protein levels typically found in dairy products [Moshtaghian et al., 2024]. Nevertheless, PBDAs may still be nutritionally limited in terms of micronutrients that are naturally abundant in dairy products, including iodine and vitamin B2, unless appropriate fortification strategies are applied [Medici et al., 2023]. From an environmental perspective, PBDAs offer substantial benefits, including reduced greenhouse gas emissions and lower land and water requirements relative to dairy production, rendering them an attractive option for consumers seeking more sustainable dietary choices [Craig et al., 2023].
A soybean yoghurt alternative offers several nutritional advantages over traditional dairy yoghurt, largely attributable to its distinctive composition and the fermentation process involved. A key benefit is its suitability for individuals with lactose intolerance and those adhering to a vegan diet, as it is entirely plant-based and free from lactose. From a nutritional perspective, a soybean yoghurt alternative is rich in proteins, vitamins, and minerals, with reported increases in B vitamins and unsaturated fatty acids compared to an unfermented soybean substrate used for its production, which are associated with cardiovascular health benefits [Mehaya et al., 2023]. Fermentation further enhances soybean nutrient bioavailability, including that of isoflavones, which have been implicated in the modulation of hyperglycaemic disorders [Langa et al., 2023]. In addition, soybean yoghurt has been shown to positively influence gut microbiota composition, an important determinant of digestive health, by promoting beneficial genera, such as Lactobacillus and Bifidobacterium [Park & Mannaa, 2025]. The presence of γ-aminobutyric acid (GABA) in certain soybean yoghurt formulations may further contribute to health-promoting effects, particularly with respect to glucose regulation and gut health [Weng et al., 2023]. Moreover, substitution of cow’s milk with a soybean substrate in yoghurt production does not appear to substantially compromise micronutrient content, as mineral and vitamin levels in the soybean yoghurt alternative are comparable to those of conventional yoghurt [Otolowo et al., 2022]. Finally, fermentation with selected lactic acid bacteria strains has been reported to enhance mineral bioavailability, which may be beneficial for bone health [Gan et al., 2023].
Co-cultivation of lactic acid bacteria (LAB) and yeast during fermentation of soybean-based substrates represents a promising strategy for enhancing the nutritional and functional properties of the final product. The combined use of LAB and yeast has been reported to improve nutrient and mineral bioavailability, as evidenced by enhanced mineral bioaccessibility and solubility, along with the degradation of phytate-mineral and protein-mineral complexes into smaller, more readily bioavailable forms during LAB-mediated fermentation [Gan et al., 2023]. In particular, fermentation of soybean substrate using Saccharomyces boulardii in combination with Lactobacillus plantarum has been shown to enhance the nutritional profile, including elevated levels of B vitamins, unsaturated fatty acids, and antioxidant activity, compared with LAB-only fermentation [Mehaya et al., 2023]. In addition, co-cultivation of selected yeasts with LAB may confer distinctive aromatic and functional attributes to soy-based fermented beverages, as demonstrated by yeast consortia that increase phenolic content and antioxidant capacity while maintaining probiotic viability [Agarbati et al., 2023]. The synergistic interactions between LAB and yeast are further supported by reports of improved carbohydrate utilisation and metabolite release, resulting in increased lactic acid production and enhanced probiotic growth [do Amaral Santos et al., 2014]. Moreover, co-cultivation strategies may mitigate undesirable off-flavours and improve sensory characteristics, as LAB are capable of producing flavour-enhancing volatile compounds [Lee et al., 2024]. These positive microbial interactions are considered critical for optimising substrate conversion and improving the nutritional and sensory quality of fermented soy products [Canon et al., 2020]. Nevertheless, the specific effects of LAB–yeast cocultivation on soybean yoghurt alternative quality parameters, including texture, protein structure, and aroma profile, remain insufficiently explored.
This study investigated the effects of co-fermentation of Levilactobacillus brevis and Saccharomyces cerevisiae on the physicochemical and structural properties of soybean-based yoghurt. By modulating initial inoculation densities, the influence of microbial interactions on gel formation, protein secondary structure, exopolysaccharide (EPS) production, and volatile organic compound profiles was systematically examined.
MATERIALS AND METHODS
Microorganism strains and culture conditions
The bacterial strain L. brevis QD-1 (NCBI accession number: PRJNA1121961) and the yeast strain S. cerevisiae (NCBI accession number: PP116084) were previously isolated from kombucha [Nguyen & Nguyen, 2024] and maintained in the microbial culture collection of the Department of Food Technology, Faculty of Applied Science and Technology (FAST), Nguyen Tat Thanh University, Ho Chi Minh City, Vietnam. L. brevis QD-1 was maintained on De Man–Rogosa–Sharpe agar (MRS; HiMedia Laboratories Pvt. Ltd., Mumbai, India), while S. cerevisiae was cultivated on yeast extract–peptone–dextrose agar (YPD; HiMedia Laboratories Pvt. Ltd., Mumbai, India). Prior to fermentation, L. brevis was propagated in MRS broth at 37°C for 18 h under anaerobic conditions, and S. cerevisiae was cultured in YPD broth at 30°C for 24 h under aerobic conditions. Preliminary growth curve experiments (data not shown) confirmed that these incubation times corresponded to late exponential phase. Following incubation, both cultures were harvested by centrifugation at 10,000×g for 5 min and washed twice with phosphate-buffered saline (PBS). The washed cell pellets were resuspended in sterile PBS and adjusted to an optical density of 1.0 measured at 600 nm (OD600), corresponding to approximately 1×10⁸ CFU/mL, to obtain standardised stock suspensions for subsequent inoculation.
Fermentation of soybean water slurry by a mixed culture of Levilactobacillus brevis QD-1 and Saccharomyces cerevisiae
High-quality, mould-free soybeans were procured from Choice L Ltd., Ho Chi Minh City, Vietnam. According to the manufacturer’s specifications, the raw soybeans contained approximately 34 g of protein, 18.4 g of lipids, 24.6 g of carbohydrates, and 4.5 g of dietary fibre per 100 g (dry weight basis). The soybeans were soaked in water for 8 h, dehulled, and ground with water at a ratio of 1:2 (g/mL). The resulting slurry was filtered through a cloth and heat-treated at 85°C until the extract reached a total solid content of 17% and an initial pH of 6.75±0.05, as measured using an Mi150 pH meter (Milwaukee Instruments, Szeged, Hungary). Following cooling to 37°C, the soybean extract was inoculated with a mixed culture of L. brevis QD-1 and S. cerevisiae at a 1:1 ratio (based on CFU counts), at final inoculation levels for each strain being 103 CFU/mL (LB+S_3), 10⁴ CFU/mL (LB+S_4), and 10⁵ CFU/mL (LB+S_5). The inoculum was added as a washed cell suspension at 1% (v/v) of the soybean extract volume. Control samples were inoculated with L. brevis QD-1 alone at the same cell densities and designated LB_3, LB_4, and LB_5, respectively. Fermentation was conducted at 37°C for 16 h in accordance with the procedure described by Nguyen et al. [2024].
Analytical methods
pH and total acidity determination
The pH was measured using a Mi150 pH meter (Milwaukee Instruments). Total titratable acidity (TTA) was determined by titrating 10 mL of the sample with 0.1 M NaOH to an endpoint of pH 8.3, using 1% (w/v) phenolphthalein as an indicator. Titratable acidity was expressed as grams of lactic acid equivalent per litre (g/L) and calculated using the following Equation (1):
where: VNaOH is the volume of 0.1 M NaOH consumed during titration (mL), Vsample is the sample volume (mL), and 0.009 is the conversion factor for a lactic acid equivalent [Nielsen, 2017].Surface morphology observation during fermentation
Surface morphology was evaluated macroscopically throughout fermentation by visual observation and photographic recording of the sample surface at different fermentation times, with particular attention to coagulation onset and the appearance of surface cracks [Nguyen et al., 2024].
Microbial density determination
Microbial density was quantified by the serial dilution plate count method. For L. brevis, appropriate dilutions were spread on MRS agar and incubated anaerobically at 37°C for 48 h. For S. cerevisiae, samples were plated on YPD agar supplemented with 1% chloramphenicol to inhibit bacterial growth and incubated aerobically at 37°C for 48 h. Colony counts were recorded and expressed as log CFU/mL [Van et al., 2023].
Exopolysaccharide quantification
Exopolysaccharides (EPS) were isolated from soybean yoghurt alternative samples using a modified method based on the protocol described by Lin & Chien [2007]. Proteins and bacterial cells were precipitated by the addition of an equal volume of 40% trichloroacetic acid, followed by centrifugation at 10,000×g for 15 min at 4°C. The resulting supernatant was mixed with an equal volume of ethanol and stored at 4°C for 24 h to allow EPS precipitation. The crude EPS was transferred into dialysis tube (10–14 kDa molecular weight cut-off) and dialysed against distilled water at 4°C for 24–48 h to remove residual medium components and other low-molecular-weight impurities. Following centrifugation under identical conditions, the EPS pellet was collected and resuspended in distilled water. The total neutral glycoside content was subsequently quantified using the phenol-sulphuric acid method, where 1 mL of the sample was mixed with 1 mL of a 5% phenol solution, followed by the rapid addition of 5 mL of concentrated sulphuric acid, incubated at room temperature for 10–30 min, and absorbance was measured at 490 nm using a glucose standard curve, with results expressed as glucose equivalents [Urshev et al., 2007].
Water-holding capacity and syneresis determination
Water-holding capacity (WHC) was determined by centrifuging 5 g of the soybean yoghurt alternative at 3,000×g for 10 min. WHC (%) was calculated using Equation (2):
where: W1 is the initial sample weight and W2 denotes the weight of the supernatant obtained after centrifugation.Syneresis was determined by storing 30 g of the soybean yoghurt alternative at 4°C for 2 h. This duration was established through preliminary trials to monitor initial whey separation while minimising confounding effects from long-term structural rearrangements. The percentage of syneresis was calculated as the mass of the expelled whey, relative to the initial sample weight [Dönmez et al., 2017].
Texture profile analysis
Texture profile analysis (TPA) was performed using a CT3 texture analyser (AMETEK Brookfield Inc., Middleboro, MA, USA) fitted with a cylindrical probe (TA4/1000; diameter 38.1 mm). The test conditions were as follows: probe-to-sample distance of 30 mm, trigger load of 7 g, and test and return speeds of 0.5 mm/s. Each sample was subjected to two consecutive compression cycles, with a total test duration of approximately 170 s and no recovery between cycles, allowing the acquisition of two positive force regions. Textural parameters, including hardness, cohesiveness, springiness, gumminess, and adhesiveness, were recorded and analysed using Text Proc CT V1.3 software (AMETEK Brookfield Inc., Middleboro, MA, USA). Hardness and gumminess were expressed in Newtons (N), converted from gram-force using the factor of 0.009807 N/g.
Sodium dodecyl sulfate–polyacrylamide gel electrophoresis
The protein composition of soybean yoghurt alternative samples was analysed by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) using 10% resolving gels [Laemmli, 1970]. Prior to electrophoresis, the samples were denatured by mixing with a sample buffer at a ratio of 4:1 (v/v), followed by heating at 100°C for 3 min and cooling for 2 min. Subsequently, 20 μL of the denatured sample were loaded into wells of a 4% stacking gel prepared with the following composition: 0.65 mL of 30% acrylamide, 1.75 mL of 0.5 mM Tris-HCl buffer (pH 6.8), 3 mL of distilled water, 100 μL of 10% SDS, 100 μL of 10% ammonium persulfate (APS), and 10 μL of N,N,N’,N’-tetramethylethylenediamine (TEMED). Electrophoresis was performed at 80 V until the proteins migrated through the stacking gel, after which the voltage was increased to 120 V for separation in the 10% resolving gel, composed of 3.3 mL of 30% acrylamide, 2.5 mL of 0.5 mM Tris-HCl buffer (pH 8.8), 4 mL of distilled water, 100 μL of 10% SDS, 100 μL of 10% APS, and 4 μL of TEMED. Following electrophoresis, the gels were stained with 0.23% Coomassie Brilliant Blue R-250 for 30 min and subsequently destained in a solution containing 10% methanol and 10% acetic acid until a clear background was achieved. Protein bands were visualised using a BioDoc-It UVP imaging system (Analytik Jena AG, Jena, Germany). Protein molecular weights were estimated by comparison with standard protein molecular weights ranging from 10 to 250 kDa (PageRuler Plus prestained protein ladder 26619, Thermo Fisher Scientific, Waltham, MA, USA). Results were expressed as qualitative band intensity comparisons.
Determination of secondary structure of protein by Fourier transform infrared spectroscopy
Lyophilised soybean yoghurt alternative samples (1 mg) were mixed with 100 mg of KBr and pressed into pellets. Fourier transform infrared (FTIR) spectroscopy was employed to characterise protein secondary structure. FTIR spectra were acquired over the wavenumber range of 400–4,000 cm–1 using an MIR Frontier FTIR spectrometer (PerkinElmer, Waltham, MA, USA) at a spectral resolution of 4 cm–1. Protein secondary structures were evaluated by focusing on the amide I region (1,600–1,700 cm–1), which is highly sensitive to backbone conformations. Prior to analysis, spectra were baseline-corrected using a second-order polynomial function. Fourier self-deconvolution (FSD) and second derivative methods were subsequently applied to enhance spectral resolution and identify underlying component peaks. Peak deconvolution and curve fitting were performed using OriginPro 2017 (Multiple Peak Fit module; OriginLab Corporation, Northampton, MA, USA), employing Gaussian functions to resolve overlapping bands. Peak positions were assigned based on established literature, corresponding to α-helix (1,650–1,658 cm–1), β-sheet (1,620–1,640 cm–1 and 1,670–1,690 cm–1), β-turn (1,660–1,680 cm–1), and random coil (1,640–1,650 cm–1) structures [Nguyen et al., 2026]. Fitting parameters were constrained within reasonable bandwidth limits (typically 10–30 cm–1), and iterative optimization was conducted until satisfactory fitting quality was achieved (R2>0.99). The relative proportion of each secondary structural element was calculated from the integrated area of individual peaks and expressed as a percentage of the total amide I band area [Yang et al., 2015].
Determination of volatile organic compounds by headspace solid-phase microextraction coupled with gas chromatography–mass spectrometry
Volatile organic compound profiles of lyophilised yoghurt alternative samples were analysed using a Nexis GC-2030 gas chromatography-mass spectrometry (GC-MS) system (Shimadzu Corporation, Kyoto, Japan) equipped with a DB-5MS capillary column (30 m × 0.25 mm × 0.25 μm) and coupled to an HS-20NX headspace sampler. Briefly, 1.0 g of each sample was placed in a 20 mL headspace glass vial, followed by the addition of 2 μL of the internal standard, 2-methyl-3-heptanone (Sigma-Aldrich, St. Louis, MO, USA). A 50/30 μm divinylbenzene/carboxen/polydimethylsiloxane (DVB/CAR/PDMS) fiber was used for extraction at 60°C for 30 min. After extraction and introduction of the headspace sample into the GC-MS system, the oven temperature programme was set as follows: initial heating to 40°C for 3 min, followed by an increase to 60°C at 10°C/min, then to 150°C at 3°C/min, and subsequently to 250°C at 20°C/min, where it was held for 5 min. Helium was used as a carrier gas at a flow rate of 1.0 mL/min, with a split ratio of 15:1. Mass spectra were acquired in a scan mode over an m/z range of 10–500. Volatile organic compounds were identified by comparison with the NIST 14L mass spectral library and semi-quantified using the internal standard, with results expressed as relative percentages (%) of the total detected volatile compounds.
Statistical analysis
All experiments were performed using independent biological replicates, and each measurement was conducted in technical triplicate. Data are presented as mean and standard deviation. Normality of the data distribution was assessed using the Shapiro–Wilk test, and homogeneity of variances was evaluated using Levene’s test. Differences among yoghurt variants were determined by one-way analysis of variance (ANOVA), followed by Tukey’s post hoc test where appropriate. Statistical significance was established at p<0.05.
RESULTS AND DISCUSSION
Changes in pH, total titratable acidity, and surface morphology during soybean yoghurt alternative production
Coagulation of soybean yoghurt alternative proteins is governed by microbial activity, acidification, and enzymatic interactions. Fermentation of soybean water slurry with L. brevis and S. cerevisiae was monitored over a 16-h period, with changes in pH and total titratable acidity presented in Figure 1. During the initial 5 h of fermentation, pH values remained relatively stable (6.12–6.29), indicating limited metabolic activity. Between the 6th and 10th h, a pronounced decline in pH was observed as a result of lactic acid accumulation (increase in total titratable acidity), with visible coagulation occurring at approximately 9 h, when pH reached 5.12–5.68. This observation is consistent with previous reports demonstrating that acidification reduces the net negative charge of soy proteins, thereby facilitating protein coagulation [Xu et al., 2019]. As pH decreases further, protein solubility diminishes, promoting the formation of larger aggregates and a denser gel network, which may lead to surface cracking [Rui et al., 2019]. In agreement with this mechanism, surface cracks were observed after 10–11 h of fermentation in all samples (Figure 2), suggesting that a fermentation duration of 9 h represents an optimal time for soybean yoghurt alternative production using L. brevis and S. cerevisiae.
Figure 1
Changes in pH and total titratable acidity during soybean yoghurt alternative production by fermentation at 37°C using different cultures. LB_3, LB_4, LB_5: fermentation with Levilactobacillus brevis QD-1 with densities of 103, 104, 105 CFU/mL, respectively (A–C). LB+S_3, LB+S_4, LB+S_5: fermentation with a mixed culture of L. brevis and Saccharomyces cerevisiae (1:1, based on CFU counts) with densities of 103, 104, 105 CFU/mL, respectively (D–F).
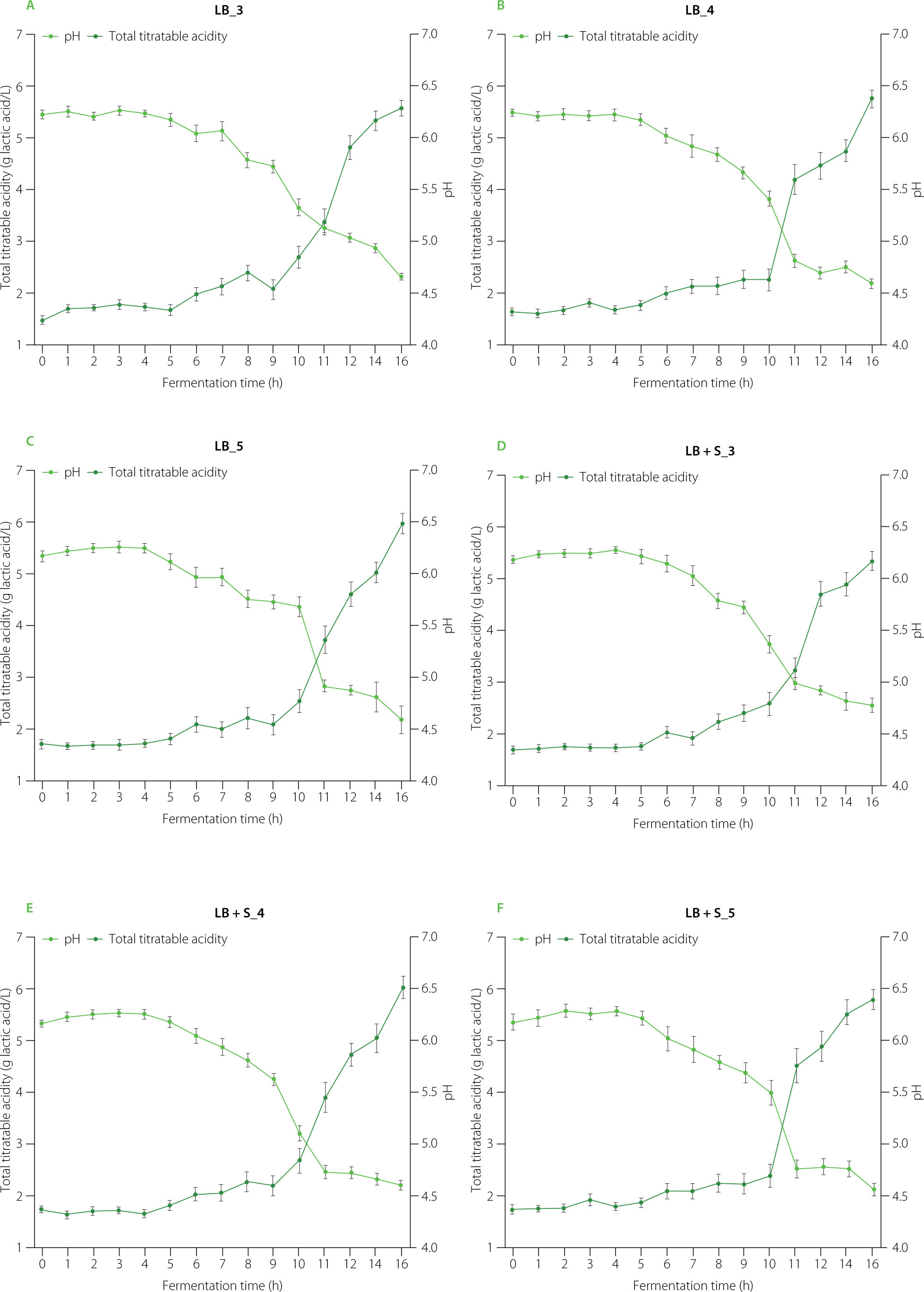
Figure 2
Surface morphology of the soybean yoghurt alternative during 16-h fermentation at 37°C using different cultures. LB_3, LB_4, LB_5: fermentation with Levilactobacillus brevis QD-1 with densities of 103, 104, 105 CFU/mL, respectively. LB+S_3, LB+S_4, LB+S_5: fermentation with a mixed culture of L. brevis and Saccharomyces cerevisiae (1:1, based on CFU counts) with densities of 103, 104, 105 CFU/mL, respectively.

It should be noted that the present study was primarily designed to provide an initial evaluation of the interactive effects between L. brevis and S. cerevisiae on soybean yoghurt alternative quality. Accordingly, the 1:1 inoculation ratio was employed as a balanced co-culture configuration to examine strain interactions under comparable initial cell densities. The incubation temperature of 37°C was selected based on the well-established performance of L. brevis in yoghurt-type fermentations [Nguyen et al., 2025]. However, both incubation temperature and inoculation ratio may influence co-fermentation dynamics between the two strains. Therefore, further studies are necessary to optimise temperature and inoculation ratio, combined with growth kinetic analysis, to gain deeper insight into each strain’s contribution to fermentation performance and product quality.
Microbial density of the soybean yoghurt alternative
Yeasts can modulate LAB populations through both synergistic and antagonistic interactions. For example, L. plantarum and S. cerevisiae can physically attach to one another, which is known as co-aggregation which promotes the formation of mixed-species biofilms. This close physical association creates a stable microenvironment that supports higher bacterial density [Furukawa et al., 2011]. Conversely, competition for nutrients may suppress LAB growth, as demonstrated by reductions in L. fermentum populations and acid production in the presence of S. cerevisiae [Gu et al., 2022]. In addition, ethanol produced by yeasts may impose osmotic and environmental stress, leading to reduced growth of certain LAB strains [Tiukova et al., 2014]. In the present study, the densities of L. brevis and S. cerevisiae were assessed at the point of soybean yoghurt alternative coagulation and were found to depend on the initial inoculum size (Table 1). Notably, no significant differences were observed in L. brevis cell density between the LB and LB+S samples at equivalent inoculation levels, indicating that the presence of S. cerevisiae did not adversely affect the growth of L. brevis under the conditions tested.
Table 1
Microbial density, exopolysaccharide content, and texture parameters of the soybean yoghurt alternative after 9-h fermentation at 37 °C using different cultures.
[i] Data expressed as the mean ± standard deviation in the same row with different lowercase letters differs significantly at p<0.05. LB_3, LB_4, LB_5: fermentation with Levilactobacillus brevis QD-1 with densities of 103, 104, 105 CFU/mL, respectively. LB+S_3, LB+S_4, LB+S_5: fermentation with a mixed culture of L. brevis and Saccharomyces cerevisiae (1:1, based on CFU counts) with densities of 103, 104, 105 CFU/mL, respectively.
Texture profile, water-holding capacity, and exopolysaccharide content of the soybean yoghurt alternative
The textural attributes of yoghurt are critical determinants of consumer acceptability, particularly for plant-based dairy alternatives. In the present study, results of texture profile analysis demonstrated that the soybean yoghurt alternative produced using a co-culture of L. brevis and S. cerevisiae exhibited significantly enhanced mechanical properties compared with the yoghurt soybean substrate fermented using L. brevis alone (Table 1). The LB+S_4 and LB+S_5 samples, inoculated at higher microbial densities (10⁴ and 10⁵ CFU/mL), recorded the highest values of selected textural parameters. These improvements were associated with elevated EPS content (201–216 mg/L), which corresponded to increased hardness (1.48–1.58 N) and gumminess (0.74–0.77 N) (Table 1). These findings are consistent with conclusions from previous studies indicating that EPS produced by lactic acid bacteria play an essential role in yoghurt structure by contributing to the formation of a dense protein-EPS network that enhances gel firmness and water retention [Akar, 2022; Brüls et al., 2024]. Notably, the co-cultivation with S. cerevisiae resulted in higher EPS levels at comparable L. brevis cell densities (Table 1). This observation aligns with the findings of Bertsch et al. [2019] who reported a 39–49% increase in EPS yield when L. rhamnosus strains (ATCC 9595, R0011, and RW-9595M) were co-cultured with S. cerevisiae. The higher gumminess of the co-cultured samples than of the monoculture treatments is consistent with the findings of Huang et al. [2020], who reported that the cofermentation of LABs and Kluyveromyces marxianus improved gel integrity and functional textural properties of goat milk. However, springiness was slightly reduced in the high-density co-culture samples (LB+S_4 and LB+S_5), and similar decreasing trends were observed for cohesiveness and adhesiveness (Table 1), indicating that although the gel matrix became denser and mechanically stronger, its elastic recovery after deformation was partially diminished. In addition, cohesiveness in the co-culture samples was not higher than in the corresponding monoculture samples at the same inoculation densities, suggesting that EPS enhancement may improve gel firmness and gumminess without necessarily increasing internal structural resilience. A similar phenomenon was described by Brüls et al. [2024], where elevated EPS levels contributed to the formation of a more compact and less elastic gel structure of yoghurt. Collectively, these findings suggest that EPS enhancement may strengthen gel structure but does not necessarily increase elasticity. Although previous studies have proposed that yeast may stimulate EPS production through interspecies metabolic interactions, such as growth factor provision or metabolite cross-feeding [Xu et al., 2025], these mechanisms were not directly examined in the present study. Therefore, further investigations incorporating quantitative analyses of yeast growth dynamics, metabolite exchange, and transcriptional regulation of EPS-related genes are necessary to elucidate the underlying mechanisms governing LAB-yeast interactions in fermentation during soybean yoghurt alternative production.
The enhanced WHC determined in the co-culture samples (Table 1) can be largely attributed to increased EPS production, which promotes the formation of a more robust gel matrix with a finer pore structure capable of retaining greater amounts of water. This interpretation is consistent with the findings of Li et al. [2014], who reported that EPS-producing Lactobacillus strains significantly improved the WHC of fermented soymilk. Similarly, Brüls et al. [2024] demonstrated that elevated EPS levels were associated with a reduced pore size and strengthened protein-EPS interactions, thereby enhancing water retention within the gel network. Despite the increase in WHC, syneresis values remained statistically comparable in all samples (70.1–77.4%), with no significant differences (p≥0.05) observed between mono-cultured and co-cultured samples (Table 1). This finding suggests that WHC and syneresis represent distinct aspects of gel behaviour: WHC reflects the capacity of the gel to retain water under applied force, whereas syneresis describes the gradual expulsion of whey under static conditions, which is more strongly influenced by gel elasticity and long-term structural stability. Moreover, although the co-cultured samples exhibited higher EPS levels, structural rearrangements within the protein matrix during storage may still have permitted water migration.
Overall, the co-fermentation of L. brevis and S. cerevisiae resulted in significant improvements in the textural quality of the soybean yoghurt alternative, which was largely attributable to enhanced EPS production. These findings indicate that cocultivation may represent a promising approach for improving the structural integrity and sensory acceptability of plant-based yoghurt alternatives.
Protein profile of the soybean yoghurt alternative
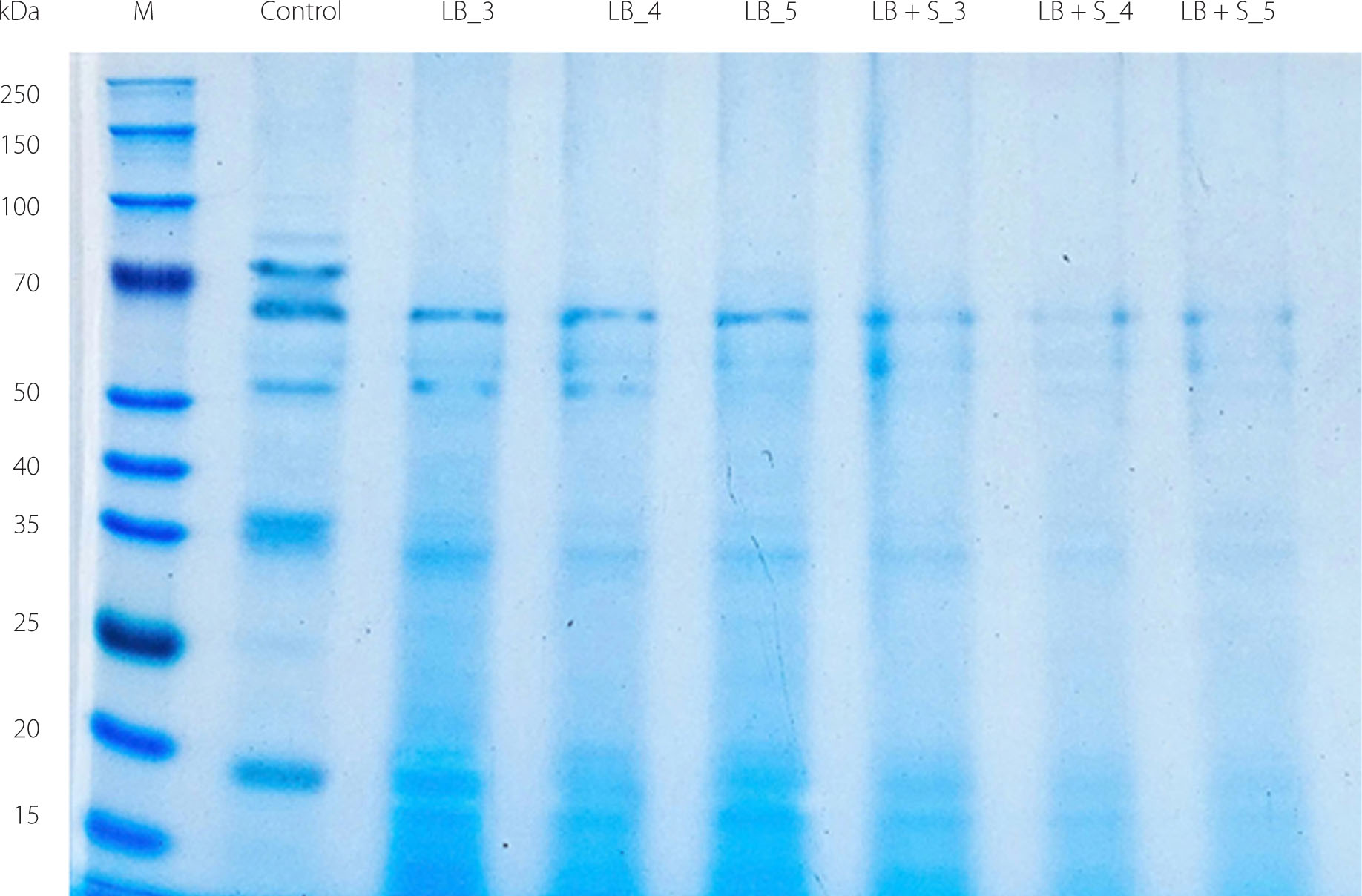
Two major storage soybean proteins are glycinin and β-conglycinin. The β-conglycinin fraction comprises three subunits, namely: α (67 kDa), α’ (71 kDa), and β (45–50 kDa) [Picariello et al., 2013], whereas glycinin consists of acidic polypeptides (40–43 kDa) and basic polypeptides (~20 kDa) [Yaklich, 2001]. Both glycinin and β-conglycinin have been reported to activate the p38/JNK/NF-κB signalling pathway, which is associated with inflammatory responses [Peng et al., 2018]. These allergenic proteins may induce hypersensitivity reactions in the skin, gastrointestinal tract, and respiratory system [Pi et al., 2021]. Notably, the β subunit of β-conglycinin exhibits highly stable tertiary and secondary structures, even following heat treatment, and is therefore relatively resistant to degradation [Pi et al., 2021]. Previous studies have demonstrated that lactic acid bacteria can degrade the α’ subunit of β-conglycinin, although the extent of hydrolysis varies among Lactobacillus species, thereby potentially reducing the allergenicity of soymilk [Aguirre et al., 2014]. Accordingly, the present study examined whether fermentation with L. brevis in combination with S. cerevisiae could reduce the levels of these allergenic proteins.
As SDS-PAGE separations show (Figure 3), L. brevis effectively degraded proteins with molecular weights of 70, 50, and 20 kDa, corresponding to the α’ and β subunits of β-conglycinin and the acidic and basic fractions of glycinin, respectively. Notably, the soybean yoghurt substrate fermented with the co-culture of L. brevis and S. cerevisiae exhibited more pronounced degradation of these protein fractions than the samples fermented with L. brevis alone. Lactic acid bacteria have been shown to produce proteases that hydrolyse soybean proteins into smaller peptides and free amino acids. This enzymatic activity modifies protein structure and contributes to changes in soybean protein gel properties during fermentation [Kieliszek et al., 2021; Ren & Li, 2022]. In addition, S. cerevisiae has been reported to produce growth-promoting factors that stimulate enzyme activity, thereby enhancing the metabolic activity of lactic acid bacteria during fermentation [Sieuwerts et al., 2018]. Such interactions may generate a synergistic effect that facilitates more efficient degradation of soy proteins. Given that β-conglycinin is recognized as one of the principal allergenic storage proteins in soybean, its structural modification during fermentation may potentially contribute to a reduction in allergenic potential [Peng et al., 2018]. Further studies incorporating targeted immunoreactivity or epitope-specific analyses would help clarify whether these structural changes are associated with modified allergenic properties.
Figure 3
Sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) separation of proteins of the soybean yoghurt alternative after 9 h of fermentation using different cultures. LB_3, LB_4, LB_5: fermentation with Levilactobacillus brevis QD-1 with densities of 103, 104, 105 CFU/mL, respectively. LB+S_3, LB+S_4, LB+S_5: fermentation with a mixed culture of L. brevis and Saccharomyces cerevisiae (1:1, based on CFU counts) with densities of 103, 104, 105 CFU/mL, respectively; M, standard proteins; Control, unfermented soybean substrate (prior to inoculation).
Changes in the protein secondary structure during soybean yoghurt alternative production
The FTIR spectra of soybean yoghurt alternative samples recorded over the wavenumber range of 400–4,000 cm–1 are presented in Figure 4A. Absorption bands observed in the regions of 3,200–3,600 cm–1 and 1,600–1,700 cm–1 correspond to the amide A and amide I bands of soy proteins, respectively, and were detected in all samples. In particular, the LB_3 sample exhibited a higher intensity of the amide A band compared to the other samples, together with the appearance of an amide IV band in the region of 530–610 cm–1. During fermentation, pH changes are known to influence the hydrogen-bond strength and the interaction patterns of amide groups within protein molecules [Liu et al., 2024]. Additionally, absorption peaks at 1,653 and 1,556 cm–1 were assigned to the stretching vibrations of carbonyl (C=O) and C–N groups, respectively, which is consistent with observations reported in recent studies on fermented soybean meal [Li et al., 2024].
Subsequently, deconvolution of the amide I band in the FTIR spectra of the soybean yoghurt alternative was performed to compare protein secondary structures based on structural components, including α-helix (1,650–1,658 cm–1), β-sheet (1,620––1,640 cm–1 and 1,670–1,690 cm–1), β-turn (1,660–1,680 cm–1), and random coil (1,640–1,650 cm–1) [Nguyen et al., 2026]. As revealed by FTIR deconvolution of the amide I band (Figure 4B), β-sheet was the dominant secondary structure in all soybean yoghurt alternative samples, ranging from 42.9% to 54.4%, which is consistent with the inherently β-sheet-rich conformation of major soy storage proteins, particularly glycinin and β-conglycinin [Yang et al., 2025]. In the L. brevis single-culture samples (LB), β-sheet content increased progressively with inoculation density, from 42.9% (LB_3) to 54.4% (LB_5), and was accompanied by a concurrent decrease in α-helix content from 25.3% to 12.3% (Figure 4B). This trend suggests that higher bacterial density promotes more extensive protein unfolding and intermolecular aggregation during fermentation, driven by accelerated acidification that destabilises native helical conformations and promotes the formation of ordered intermolecular hydrogen-bonded β-sheet networks [Liu et al., 2022a]. The reduction in α-helix content at higher inoculation densities is indicative of greater disruption of the compact globular structure of soy proteins [Ai et al., 2019], which is consistent with enhanced proteolytic activity and lower pH environments produced by denser L. brevis populations.
Figure 4
(A) Fourier transform infrared (FTIR) spectra with focused bands of amide I and (B) secondary structure contents of protein in the soybean yoghurt alternative after 9 h of fermentation using different cultures: LB_3, LB_4, LB_5: fermentation with Levilactobacillus brevis QD-1 with densities of 103, 104, 105 CFU/mL, respectively. LB+S_3, LB+S_4, LB+S_5: fermentation with a mixed culture of L. brevis and Saccharomyces cerevisiae (1:1, based on CFU counts) with densities of 103, 104, 105 CFU/mL, respectively.
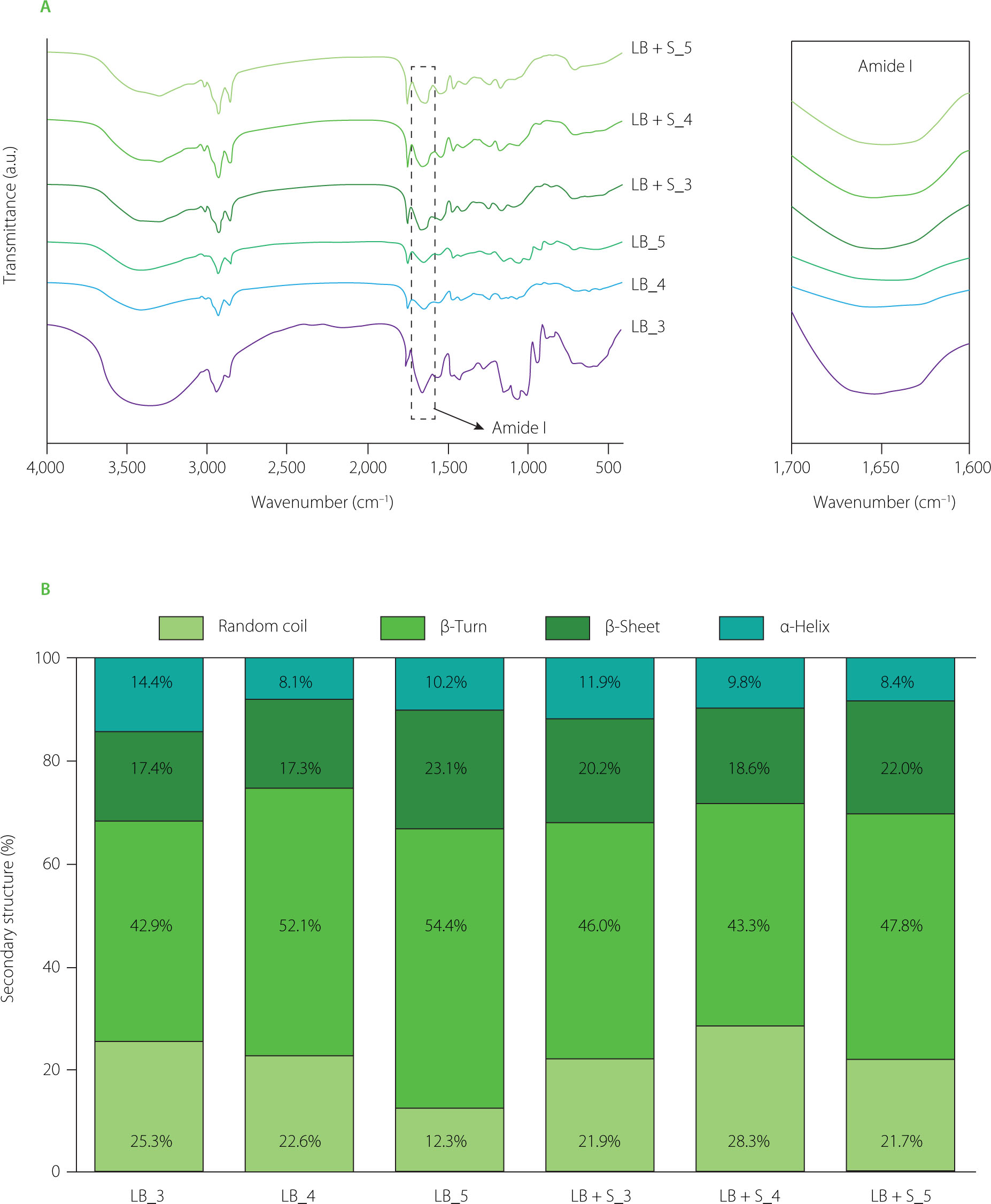
In the mixed-culture samples (LB+S), the overall structural profile was broadly similar to the single-culture counterparts, with β-sheet remaining the predominant structure (43.3–47.8%). However, α-helix content was generally higher in LB+S samples compared to their LB equivalents at the same inoculation density, most notably at the 10⁴ CFU/mL level (LB+S_4: 28.3% vs. LB_4: 22.6%) (Figure 4B), suggesting that the co-fermentation with S. cerevisiae partially moderates protein denaturation. This may be attributed to the metabolic activity of S. cerevisiae, which produces CO2 and ethanol alongside organic acids, potentially creating a different physicochemical environment that slows the complete unfolding of helical domains. Furthermore, random coil content tended to be lower in LB+S samples than in the LB samples, implying that the mixed fermentation system maintains a more ordered protein conformation, possibly through the combined effect of moderate acidification and yeast-derived compounds that interact with the protein matrix [Liu et al., 2022b]. In turn, β-turn content remained relatively stable (17.3–23.1%) in both fermentation systems (Figure 4B), indicating that these structural elements, which serve as flexible connectors between β-strands, are less sensitive to fermentation conditions than the α-helix or random coil fractions. These findings demonstrate that both inoculation density and culture composition significantly modulate the protein conformational changes during soybean yoghurtsubstrate fermentation, with implications for gel texture, water-holding capacity, and digestibility of the final product. The representative deconvoluted spectra and corresponding quantitative data are provided in Figure S1 and Table S1 (Supplementary Materials), respectively.
Volatile organic compounds of the soybean yoghurt alternative
The volatile organic compound profiles of the soybean yoghurt produced with L. brevis QD-1 at different inoculation densities, with and without co-cultivation with S. cerevisiae, are summarised in Table 2. Ethanol was the predominant volatile organic compound detected in all six samples, accounting for 91.02–92.83% of the total volatile organic compounds in the LB sample and 86.87–93.51% in the LB+S sample, followed by acetic acid (4.30––4.65% in the LB sample and 4.76–5.42% in the LB+S samples) and 2,3-butanediol (0.95–2.35% in the LB samples and 0.37––9.46% in the LB+S samples). L. brevis, as a heterofermentative lactic acid bacterium, possesses a metabolic framework that markedly influences the production of ethanol and acetic acid during fermentation. Its metabolic versatility enables the utilisation of various carbohydrates, including glucose and fructose, primarily via the phosphoketolase pathway, which underpins its heterofermentative metabolism. This pathway yields lactic acid alongside ethanol and acetic acid as major by-products, the latter two potentially constraining the efficiency of lactic acid production [Guo et al., 2014].
Table 2
Volatile organic compound profile of the soybean yoghurt alternative after 9-h fermentation at 37°C using different cultures.
[i] Data are mean ± standard deviation (n=3). Means in the same row with different lowercase letters differ significantly (p<0.05). LB_3, LB_4, LB_5: fermentation with Levilactobacillus brevis QD-1 with densities of 103, 104, 105 CFU/mL, respectively. LB+S_3, LB+S_4, LB+S_5: fermentation with a mixed culture of L. brevis and Saccharomyces cerevisiae (1:1, based on CFU counts) with densities of 103, 104, 105 CFU/mL, respectively. RT, retention time; RI, retention index; MW, molecular weight; nd, not detected.
However, co-cultivation with S. cerevisiae, particularly at an inoculation density of 10⁵ CFU/mL, markedly altered the volatile organic compound profile of the soybean yoghurt alternative. This shift was characterised by a smaller relative proportion of ethanol (69.55%) alongside increased relative levels of acetic acid (15.42%), 2,3-butanediol (9.46%), and phenethyl alcohol (1.87%). Elevated levels of polyols and aldehydes were recorder during high-density co-fermentation, which has been attributed to the upregulation of genes involved in carbohydrate and amino acid metabolism [Zhu et al., 2023]. Saccharomyces species primarily ferment sugars into ethanol and carbon dioxide and, while they can contribute to flavour development and textural modification, they do not substantially enhance isoflavone bioavailability nor generate the same diversity of bioactive compounds as LAB [Sharma et al., 2023]. The aroma profile generated during fermentation is strongly influenced by both the amino acid composition of the substrate and the yeast strain employed, as distinct strains exhibit unique metabolic capacities that govern the synthesis of volatile organic compounds [Scott et al., 2021]. In particular, the availability of specific amino acids can direct aroma formation, with methionine metabolism yielding methionol and its acetate ester, compounds that contribute to savoury flavour notes [Etschmann et al., 2008]. In contrast, LAB, including L. plantarum and L. rhamnosus, are well recognised for their capacity to biotransform isoflavones into more bioavailable aglycones, such as daidzein, genistein, glycitein, and further into metabolites with enhanced biological activity, including equol, dihydrodaidzein, and dihydrogenistein [Langa et al., 2023]. These transformations are mediated by LAB-derived β-glucosidase activity, which promotes the release of bound phenolic compounds including isoflavones, thereby increasing the bioactivity of the fermented product [Zhang et al., 2023]. In addition, LAB contributes to the generation of bioactive peptides with antioxidant properties through the proteolysis of soybean proteins, such as glycinin and β-conglycinin, which are abundant in germinated soymilk [Fadlillah et al., 2025]. Comparable effects have also been reported in kombucha fermentation systems, where the incorporation of LAB enhanced flavour complexity through increased production of alcohols and esters [Wang et al., 2024]. Overall, these findings indicate that the metabolic interactions between LAB and yeast can promote the formation of a broader and more balanced volatile organic compound profile, which is critical for improving the sensory quality of fermented foods and beverages. However, volatile organic compounds were analysed semi-quantitatively and expressed as relative percentages rather than absolute concentrations; therefore, the data reflect shifts in aroma composition rather than definitive differences in ethanol yield. A smaller relative ethanol proportion does not necessarily indicate reduced absolute production but may instead result from increased synthesis of other volatiles that affect proportional distribution. Further investigation incorporating absolute ethanol quantification, residual sugar analysis, and CO2 measurement is required to clarify the metabolic contribution of S. cerevisiae to ethanol dynamics in yoghurt fermentation during soybean yoghurt alternative production.
CONCLUSIONS
In summary, this study demonstrates that the co-inoculation of L. brevis QD-1 with S. cerevisiae produced a soybean-based yoghurt alternative with improved textural properties associated with higher EPS levels, more extensive degradation of major soy storage proteins (β-conglycinin and glycinin), and favourable modifications in protein secondary structure compared with the L. brevis monoculture. Volatile profiles also showed shifts in the relative abundances of key compounds. The viability of L. brevis was not negatively affected by the presence of the yeast. These outcomes indicate that S. cerevisiae can serve as a supportive costarter culture capable of modulating selected quality attributes of the soy yoghurt alternative produced by L. brevis-fermentation, where acidification and primary gel formation are driven by the lactic acid bacterium. The study results support the potential of this co-culture strategy for the development of high-quality vegan yoghurt alternatives. However, further studies are needed to clarify the respective contributions of each strain, optimise fermentation conditions, and evaluate sensory properties and consumer acceptability. Such work will help to better define the potential of LAB–yeast co-fermentation for developing plantbased yoghurt alternatives.